マムシの熱穴は、吻の深いポケットに膜が張られた構造になっています。 膜の裏側には空気で満たされた室があり、膜の両側で空気と接触している。 ピット膜は非常に血管が多く、三叉神経の終末塊から形成される多数の熱感受性受容体(terminal nerve masses, TNM)で強く神経支配されている。 したがって受容体は個別の細胞ではなく、三叉神経自体の一部なのです。 ボアやニシキヘビに見られる唇孔は浮遊膜を欠き、より単純に膜で覆われた孔からなり、同様に神経と血管が通っているが、血管の形態はこれらのヘビとクロテンで異なっている。 血管系の目的は、受容体末端に酸素を供給することに加えて、刺激による熱放射で温められた受容体を速やかに熱中性状態に冷却することである。 もしこの血管系がなければ、受容体は暖かい刺激にさらされた後でも暖かい状態のままであり、刺激を取り除いた後でも動物に残像を見せることになる。
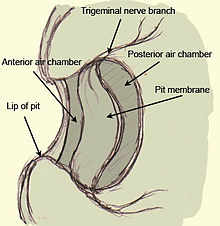
Crotaline pit organの図。
NeuroanatomyEdit
いずれの場合も、顔面ピットは三叉神経によって神経支配されています。 クロタリンでは、pit器官からの情報は外側下行三叉神経路を経て髄質の網目状核に中継される。 そこから対側の視蓋に中継される。 ボアやパイソンでは、唇孔からの情報は、側方下行三叉路を経由して、網様体路核を迂回して対側の視蓋に直接送られる
これらの赤外線の手がかりを最終的に処理するのは、脳の視蓋である。 この部分は他の感覚情報、特に視神経刺激も受け取るが、運動、固有感覚、聴覚も受け取る。 視覚や赤外線の刺激に単独で反応する神経細胞もあれば、視覚と赤外線の複合刺激に強く反応する神経細胞もあり、また視覚と赤外線の複合刺激のみに反応する神経細胞もある。 また、一方向の動きを感知するように調整されている神経細胞もあるようだ。 ヘビの視覚と赤外線の世界地図は、視蓋で重ね合わされていることが分かっている。 この結合された情報は、視蓋を経由して前脳に伝えられます。
ピット器官の神経線維は、非常に低い割合で常に発火しています。 中性温度範囲にある物体は発火率を変えない。中性温度範囲は器官の受容野にあるすべての物体の平均熱放射によって決定される。 ある閾値以上の熱放射は神経繊維の温度上昇を引き起こし、その結果、神経が刺激され、それに続いて発火し、温度上昇により発火率が上昇する。 神経線維の感度は>0.001℃とされている。
ピット器官は繰り返しの刺激に適応し、適応した刺激を取り除くと、反対方向の変動が生じる。 例えば、蛇の前に暖かいものを置くと、最初は発火率が上がるが、しばらくすると暖かいものに順応し、ピットオルガンの神経の発火率は元に戻る。 その後、その暖かい物体を取り除くと、今度は自分がいた空間を寒いと認識し、物体が取り除かれたことに適応するまで発火率が低下する。 適応の潜伏期間は約50~150msである。
顔面ピットは実際にはピンホールカメラと同じ光学原理で熱放射を視覚化しており、熱放射の発生源の位置は、熱ピットの膜上の放射の位置によって決定される。 しかし、フェイシャルピットが見ている熱画像をコンピューター解析で可視化した研究によると、解像度が極めて低いことが指摘されている。 ピットの開口部の大きさから、小さな温熱物体の解像度が悪く、ピットの小ささとそれに伴う熱伝導の悪さと相まって、生成される画像は極めて低い解像度とコントラストとなるのです。 外側下行三叉路で画像の焦点合わせと鮮鋭化が行われることが知られているが、眼窩で行われる視覚と赤外線の統合も画像の鮮鋭化に役立っている可能性がある。
分子機構編集
赤外線を検出するにもかかわらず、赤外線検出機構は視細胞とは似て非なるもので、視細胞が光化学反応によって光を検出するのに対し、蛇の孔のタンパク質は温度感受性イオンチャンネルであるTRPV1という一過性の受容電位チャネルの一種であった。 光に対する化学反応ではなく、ピット器官の加温を伴うメカニズムで赤外線信号を感知している。 その構造と機能は、ボロメーターと呼ばれる生物版温感器に似ている。 これは、ピット膜が薄いため、入射した赤外線が迅速かつ正確に所定のイオンチャネルを温めて神経インパルスを誘発すること、また、イオンチャネルを迅速に元の温度状態に冷却するためにピット膜が血管化されていることと矛盾しない。 このメカニズムの分子的前駆体は他のヘビにも見られるが、このタンパク質の発現量ははるかに低く、熱に対する感受性もはるかに低い
。