DNA-replikation, også kendt som semi-konserverende replikation, er den proces, hvorved DNA i det væsentlige fordobles. Det er en vigtig proces, der finder sted i den delende celle.
I denne artikel skal vi kort se på DNA’s struktur, på de præcise trin, der er involveret i replikation af DNA (initiering, forlængelse og afslutning), og de kliniske konsekvenser, der kan opstå, når det går galt.
DNA’s struktur
DNA består af millioner af nukleotider. Disse er molekyler, der består af et deoxyribose-sukker med en fosfat og en base (eller nukleobase) knyttet til det. Disse nukleotider er knyttet til hinanden i strenge via phosphodiesterbindinger for at danne en “sukker-fosfat-rygge”. Den dannede binding er mellem det tredje kulstofatom på deoxyribose-sukkeret på det ene nukleotid (fremover kaldet 3′) og det femte kulstofatom på et andet sukker på det næste nukleotid (kaldet 5′).
N.B.: 3′ udtales “tre primtal” og 5′ udtales “fem primtal”.
Der er to strenge, der løber i modsatte eller antiparallelle retninger til hinanden. De er knyttet til hinanden i hele strengen gennem baserne på hvert nukleotid i hele dens længde. Der er 4 forskellige baser forbundet med DNA; Cytosin, Guanin, Adenin og Thymin. I normale DNA-strenge binder cytosin sig til guanin, og adenin binder sig til thymin. De to strenge danner sammen en dobbeltspiral.
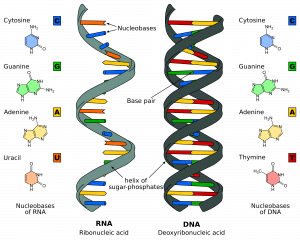 Fig 1.0 – Strukturen af RNA og DNA
Fig 1.0 – Strukturen af RNA og DNADNA-replikationens faser
DNA-replikation kan tænkes i tre faser; initiering, forlængelse og afslutning
Initiering
DNA-syntesen initieres på bestemte punkter i DNA-strengen, der er kendt som “origins”, som er specifikke kodningsregioner. Disse origins er mål for initiatorproteiner, som derefter rekrutterer flere proteiner, der hjælper med replikationsprocessen og danner et replikationskompleks omkring DNA-originet. Der er flere oprindelsessteder, og når replikationen af DNA begynder, kaldes disse steder for replikationsgafler.
I replikationskomplekset findes enzymet DNA-helicase, som afvikler dobbelthelixen og blotlægger hver af de to strenge, så de kan bruges som skabelon til replikation. Det gør dette ved at hydrolyse den ATP, der er brugt til at danne bindingerne mellem nukleobaserne, og dermed bryde den binding, der holder de to strenge sammen.
DNA-primase er et andet enzym, der er vigtigt i DNA-replikationen. Det syntetiserer en lille RNA-primer, der fungerer som en “kickstarter” for DNA-polymerase. DNA-polymerase er det enzym, der i sidste ende er ansvarlig for dannelsen og udvidelsen af de nye DNA-strenge.
Forlængelse
Når DNA-polymerasen har sat sig fast på de oprindelige, udrevne to DNA-strenge (dvs. skabelonstrengene), kan den begynde at syntetisere det nye DNA, så det passer til skabelonerne. Det er vigtigt at bemærke, at DNA-polymerase kun er i stand til at forlænge primeren ved at tilføje frie nukleotider i 3′-enden.
Den ene af skabelonerne læses i en 3′- til 5′-retning, hvilket betyder, at den nye streng vil blive dannet i en 5′- til 3′-retning. Denne nyligt dannede streng betegnes som den ledende streng. Langs denne streng behøver DNA-primase kun at syntetisere en RNA-primer én gang, i begyndelsen, for at igangsætte DNA-polymerase. Dette skyldes, at DNA Polymerase er i stand til at forlænge den nye DNA-streng ved at læse skabelonen fra 3′ til 5′ og syntetisere i en 5′ til 3′-retning som nævnt ovenfor.
Den anden skabelonstreng (den lagging-streng) er imidlertid antiparallel og læses derfor i en 5′ til 3′-retning. Kontinuerlig DNA-syntese, som i den førende streng, ville skulle foregå i 3′-til-5′-retningen, hvilket er umuligt, da vi ikke kan tilføje baser til 5′-enden. I stedet tilføjes RNA-præmier til de nyligt eksponerede baser på den bagvedliggende streng, efterhånden som spiralen rulles ud, og DNA-syntesen sker i fragmenter, men stadig i 5′-til-3′-retningen som tidligere. Disse fragmenter er kendt som Okazaki-fragmenter.
Terminering
Processen med at udvide de nye DNA-strenge fortsætter, indtil der enten ikke er mere DNA-skabelon tilbage at replikere (dvs. for enden af kromosomet), eller to replikationsgafler mødes og efterfølgende termineres. Mødet mellem to replikationsgafler er ikke reguleret og sker tilfældigt langs kromosomets forløb.
Når DNA-syntesen er afsluttet, er det vigtigt, at de nyligt syntetiserede strenge er bundet og stabiliseret. Med hensyn til den efterslæbende streng er der brug for to enzymer for at opnå dette; RNAase H fjerner RNA-primeren, der er i begyndelsen af hvert Okazaki-fragment, og DNA-ligase sammenføjer fragmenterne til én komplet streng.
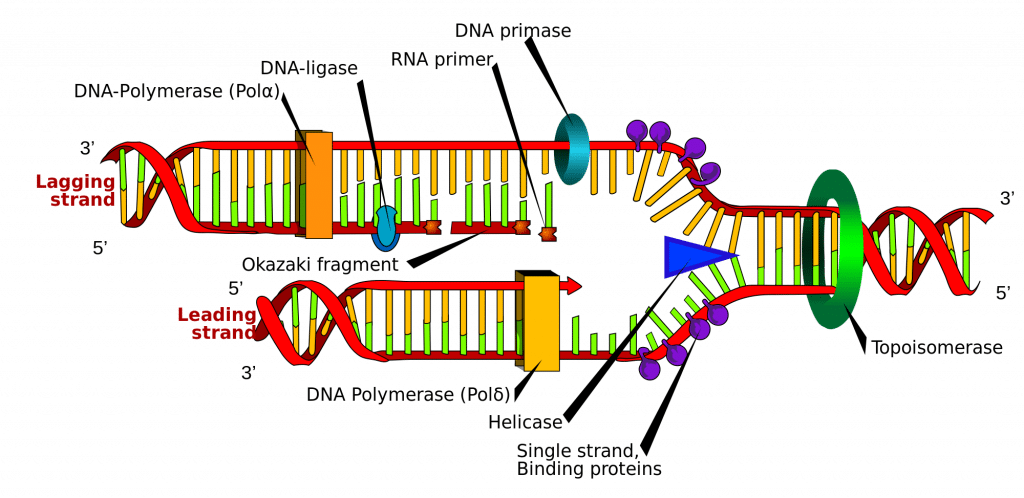 Fig. 2.0 – Diagrammatisk fremstilling af DNA-replikation
Fig. 2.0 – Diagrammatisk fremstilling af DNA-replikation
Klinisk relevans – seglcelleanæmi
Seglcelleanæmi er en autosomal recessiv tilstand, der skyldes en enkelt basesubstitution, hvor kun én base udskiftes med en anden. I nogle tilfælde kan dette resultere i en “tavs mutation”, hvor det samlede gen ikke påvirkes, men i sygdomme som f.eks. seglcelleanæmi resulterer det i, at strengen koder for et andet protein.
I dette tilfælde er en adeninbase byttet ud med en thyminbase i et af de gener, der koder for hæmoglobin; dette resulterer i, at glutaminsyre erstattes med valin. Når dette transskriberes til en polypeptidkæde, ændres dens egenskaber radikalt, da glutaminsyre er hydrofilt, mens valin er hydrofobt. Denne hydrofobiske region resulterer i, at hæmoglobin har en unormal struktur, som kan forårsage blokering af kapillærer, der fører til iskæmi og potentielt nekrose af væv og organer – dette er kendt som en vaso-okklusiv krise.
Disse kriser håndteres typisk med en række forskellige smertestillende lægemidler, herunder opioider og NSAID’er afhængigt af sværhedsgraden. I nødstilfælde kan det være nødvendigt med transfusioner af røde blodlegemer, f.eks. hvis blokaden opstår i lungerne.
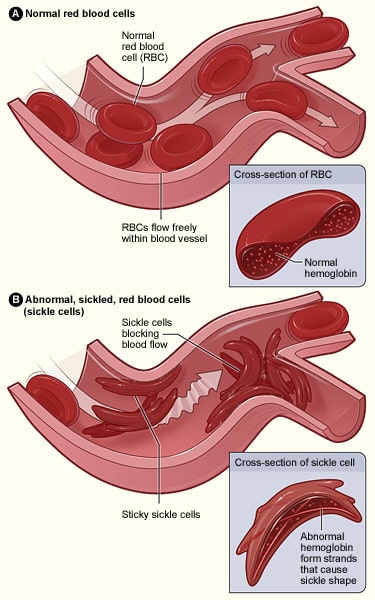 Fig 3.0 – Forskellen i struktur mellem normale røde blodlegemer og dem, der er ramt af seglcellesygdom.
Fig 3.0 – Forskellen i struktur mellem normale røde blodlegemer og dem, der er ramt af seglcellesygdom.