DNA-replikation, även känd som semi-konservativ replikation, är den process genom vilken DNA i princip dubbleras. Det är en viktig process som äger rum i den delande cellen.
I den här artikeln ska vi kortfattat titta på DNA:s struktur, på de exakta stegen vid replikering av DNA (initiering, förlängning och avslutande) och de kliniska konsekvenser som kan uppstå när detta går fel.
DNA:s struktur
DNA består av miljoner nukleotider. Dessa är molekyler som består av ett deoxyribosocker med en fosfat och en bas (eller nukleobas) knutna till det. Dessa nukleotider är knutna till varandra i strängar via fosfodiesterbindningar för att bilda en ”socker-fosfatryggrad”. Den bindning som bildas är mellan den tredje kolatomen på deoxyribosesockret i en nukleotid (hädanefter kallad 3′) och den femte kolatomen på ett annat socker i nästa nukleotid (kallad 5′).
N.B.: 3′ uttalas ”three prime” (tre primörer) och 5′ uttalas ”five prime” (fem primörer).
Det finns två strängar som löper i motsatt eller antiparallellell riktning till varandra. De är fästa vid varandra i hela strängens längd genom baserna på varje nukleotid. Det finns fyra olika baser associerade med DNA; cytosin, guanin, adenin och tymin. I normala DNA-strängar binder cytosin till guanin och adenin till tymin. De två strängarna bildar tillsammans en dubbelspiral.
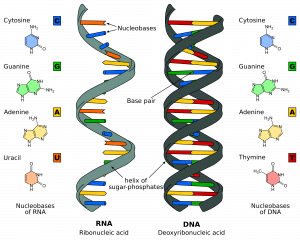 Fig 1.0 – RNA:s och DNA:s struktur
Fig 1.0 – RNA:s och DNA:s strukturDNA-replikationens stadier
DNA-replikation kan betraktas i tre stadier; Initiering, förlängning, avslutande
Initiering
DNA-syntesen initieras vid särskilda punkter i DNA-strängen, så kallade ”origins”, som är specifika kodningsområden. Dessa origins är måltavlor för initiatorproteiner, som sedan rekryterar fler proteiner som hjälper till med replikationsprocessen och bildar ett replikationskomplex runt DNA-originet. Det finns flera ursprungsplatser, och när replikationen av DNA börjar kallas dessa platser för replikationsgafflar.
Inom replikationskomplexet finns enzymet DNA-helicas, som avvecklar dubbelhelixen och frilägger var och en av de två strängarna, så att de kan användas som en mall för replikation. Det gör detta genom att hydrolysa den ATP som används för att bilda bindningarna mellan nukleobaserna och därmed bryta den bindning som håller ihop de två strängarna.
DNA-primas är ett annat enzym som är viktigt vid DNA-replikation. Det syntetiserar en liten RNA-primer, som fungerar som en ”kick-starter” för DNA-polymeras. DNA-polymeras är det enzym som i slutändan ansvarar för skapandet och utbyggnaden av de nya DNA-strängarna.
Förlängning
När DNA-polymeraset väl har fäst sig vid de ursprungliga, uppluckrade två DNA-strängarna (dvs. mallsträngarna) kan det börja syntetisera det nya DNA:t så att det matchar mallarna. Det är viktigt att notera att DNA-polymeras endast kan förlänga primern genom att lägga till fria nukleotider i 3′-ändan.
En av mallarna avläses i en 3′- till 5′-riktning, vilket innebär att den nya strängen kommer att bildas i en 5′- till 3′-riktning. Denna nybildade sträng kallas den ledande strängen. Längs denna sträng behöver DNA-primaset bara syntetisera en RNA-primer en gång, i början, för att initiera DNA-polymeraset. Detta beror på att DNA-polymeras kan förlänga den nya DNA-strängen genom att läsa av mallen 3′ till 5′ och syntetisera i en 5′ till 3′-riktning enligt ovan.
Den andra mallensträngen (den eftersläpande strängen) är dock antiparallellel och läses därför i en 5′ till 3′-riktning. Kontinuerlig DNA-syntes, som i den ledande strängen, skulle behöva ske i 3′- till 5′-riktningen, vilket är omöjligt eftersom vi inte kan lägga till baser i 5′-ändan. I stället läggs RNA-primers till de nyligen exponerade baserna på den eftersläpande strängen när spiralen rullas ut, och DNA-syntesen sker i fragment, men fortfarande i 5′- till 3′-riktningen som tidigare. Dessa fragment kallas Okazaki-fragment.
Terminering
Processen med att expandera de nya DNA-strängarna fortsätter tills det antingen inte finns någon mer DNA-mall kvar att replikera (dvs. i slutet av kromosomen), eller så möts två replikationsgafflar och avslutas därefter. Mötet mellan två replikationsgafflar är inte reglerat och sker slumpmässigt längs kromosomens förlopp.
När DNA-syntesen är avslutad är det viktigt att de nysyntetiserade strängarna binds och stabiliseras. När det gäller den eftersläpande strängen behövs två enzymer för att uppnå detta; RNAas H tar bort RNA-primern som finns i början av varje Okazaki-fragment, och DNA-ligas förenar fragmenten för att skapa en komplett sträng.
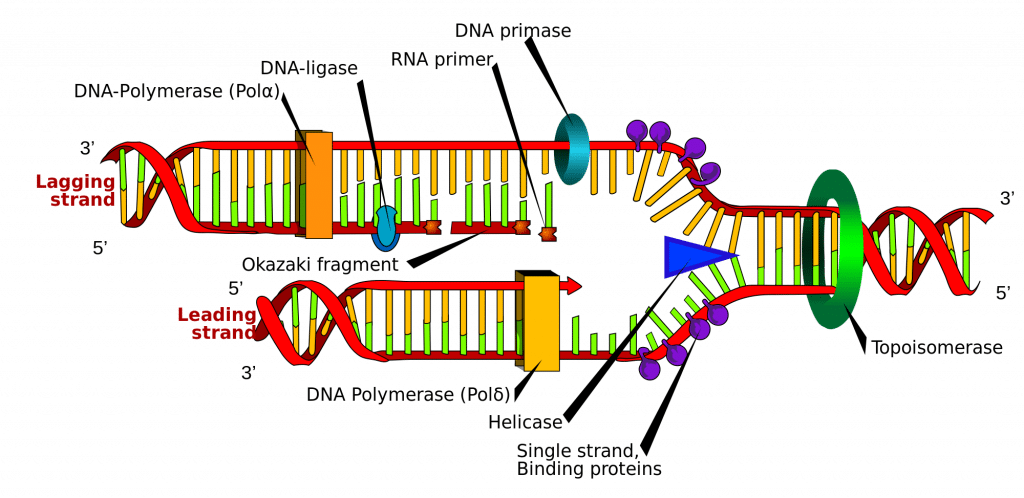 Fig 2.0 – Schematisk representation av DNA-replikation
Fig 2.0 – Schematisk representation av DNA-replikation
Klinisk relevans – Sicklecellanemi
Sicklecellanemi är ett autosomalt recessivt tillstånd som orsakas av en enkelbasersättning, där endast en bas byts ut mot en annan. I vissa fall kan detta resultera i en ”tyst mutation” där den totala genen inte påverkas, men i sjukdomar som sicklecellanemi resulterar det i att strängen kodar för ett annat protein.
I detta fall byts en adeninbas ut mot en tyminbas i en av de gener som kodar för hemoglobin; detta resulterar i att glutaminsyra ersätts med valin. När detta transkriberas till en polypeptidkedja ändras dess egenskaper radikalt eftersom glutaminsyra är hydrofil, medan valin är hydrofob. Denna hydrofoba region resulterar i att hemoglobin har en onormal struktur som kan orsaka blockering av kapillärer vilket leder till ischemi och potentiell nekros av vävnader och organ – detta är känt som en vaso-okclusiv kris.
Dessa kriser hanteras vanligen med en mängd olika smärtstillande läkemedel, inklusive opioider och NSAID beroende på svårighetsgraden. Transfusioner av röda blodkroppar kan krävas i nödsituationer, till exempel om blockeringen sker i lungorna.
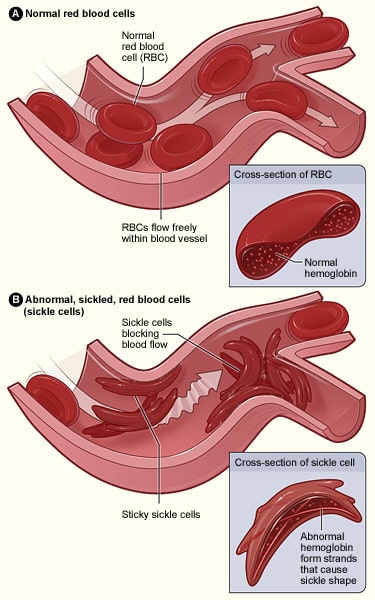 Fig 3.0 – Skillnaden i struktur mellan normala röda blodkroppar och de som är drabbade av sicklecellsjukdom.
Fig 3.0 – Skillnaden i struktur mellan normala röda blodkroppar och de som är drabbade av sicklecellsjukdom.