La réplication de l’ADN, également appelée réplication semi-conservative, est le processus par lequel l’ADN est essentiellement doublé. C’est un processus important qui a lieu au sein de la cellule en division.
Dans cet article, nous examinerons brièvement la structure de l’ADN, les étapes précises de la réplication de l’ADN (initiation, élongation et terminaison) et les conséquences cliniques qui peuvent survenir lorsque cela se passe mal.
Structure de l’ADN
L’ADN est constitué de millions de nucléotides. Ce sont des molécules composées d’un sucre désoxyribose, auquel sont attachés un phosphate et une base (ou nucléobase). Ces nucléotides sont attachés les uns aux autres en brins par des liaisons phosphodiester pour former un « squelette sucre-phosphate ». La liaison formée est entre le troisième atome de carbone sur le sucre désoxyribose d’un nucléotide (désormais appelé 3′) et le cinquième atome de carbone d’un autre sucre sur le nucléotide suivant (appelé 5′).
N.B : 3′ se prononce « trois prime » et 5′ se prononce « cinq prime ».
Il y a deux brins qui s’étendent dans des directions opposées ou antiparallèles l’un à l’autre. Ils sont attachés l’un à l’autre sur toute la longueur du brin par les bases de chaque nucléotide. Il existe 4 bases différentes associées à l’ADN : la cytosine, la guanine, l’adénine et la thymine. Dans les brins d’ADN normaux, la cytosine se lie à la guanine et l’adénine se lie à la thymine. Les deux brins forment ensemble une double hélice.
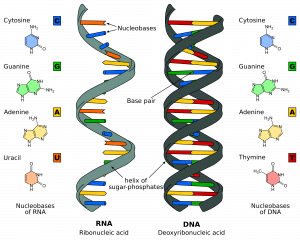 Fig 1.0 – La structure de l’ARN et de l’ADN
Fig 1.0 – La structure de l’ARN et de l’ADNÉtapes de la réplication de l’ADN
La réplication de l’ADN peut être envisagée en trois étapes ; Initiation, Élongation, Terminaison
Initiation
La synthèse de l’ADN est initiée à des points particuliers du brin d’ADN appelés » origines « , qui sont des régions codantes spécifiques. Ces origines sont ciblées par des protéines initiatrices, qui recrutent ensuite d’autres protéines qui contribuent au processus de réplication, formant un complexe de réplication autour de l’origine de l’ADN. Il existe plusieurs sites d’origine, et lorsque la réplication de l’ADN commence, ces sites sont appelés fourches de réplication.
A l’intérieur du complexe de réplication se trouve l’enzyme ADN Helicase, qui déroule la double hélice et expose chacun des deux brins, afin qu’ils puissent être utilisés comme modèle pour la réplication. Elle le fait en hydrolysant l’ATP utilisé pour former les liaisons entre les nucléobases, brisant ainsi la liaison qui maintient les deux brins ensemble.
L’ADN Primase est une autre enzyme importante dans la réplication de l’ADN. Elle synthétise une petite amorce d’ARN, qui agit comme un » coup de pouce » pour l’ADN polymérase. L’ADN polymérase est l’enzyme qui est finalement responsable de la création et de l’expansion des nouveaux brins d’ADN.
Elongation
Une fois que l’ADN polymérase s’est attachée aux deux brins d’ADN originaux et dézippés (c’est-à-dire les brins modèles), elle est capable de commencer à synthétiser le nouvel ADN pour correspondre aux modèles. Il est essentiel de noter que l’ADN polymérase n’est capable d’étendre l’amorce qu’en ajoutant des nucléotides libres à l’extrémité 3′.
L’une des matrices est lue dans le sens 3′ vers 5′, ce qui signifie que le nouveau brin sera formé dans le sens 5′ vers 3′. Ce brin nouvellement formé est appelé le brin principal. Le long de ce brin, l’ADN polymérase n’a besoin de synthétiser une amorce d’ARN qu’une seule fois, au début, pour initier l’ADN polymérase. Cela est dû au fait que l’ADN polymérase est capable d’étendre le nouveau brin d’ADN en lisant la matrice de 3′ en 5′, synthétisant dans une direction de 5′ en 3′ comme indiqué ci-dessus.
Cependant, l’autre brin de la matrice (le brin retardé) est antiparallèle, et est donc lu dans une direction de 5′ en 3′. La synthèse continue de l’ADN, comme dans le brin menant, devrait se faire dans le sens 3′ vers 5′, ce qui est impossible car nous ne pouvons pas ajouter de bases à l’extrémité 5′. Au lieu de cela, au fur et à mesure que l’hélice se déroule, des amorces d’ARN sont ajoutées aux bases nouvellement exposées sur le brin de fuite et la synthèse de l’ADN se produit par fragments, mais toujours dans le sens 5′ vers 3′ comme auparavant. Ces fragments sont connus sous le nom de fragments d’Okazaki.
Termination
Le processus d’expansion des nouveaux brins d’ADN se poursuit jusqu’à ce qu’il ne reste plus de matrice d’ADN à répliquer (c’est-à-dire à l’extrémité du chromosome), ou que deux fourches de réplication se rencontrent et se terminent ensuite. La rencontre de deux fourches de réplication n’est pas régulée et se produit au hasard le long du parcours du chromosome.
Une fois la synthèse de l’ADN terminée, il est important que les brins nouvellement synthétisés soient liés et stabilisés. En ce qui concerne le brin retardé, deux enzymes sont nécessaires pour y parvenir ; l’ARNase H élimine l’amorce d’ARN qui se trouve au début de chaque fragment d’Okazaki, et l’ADN Ligase réunit les fragments pour créer un brin complet.
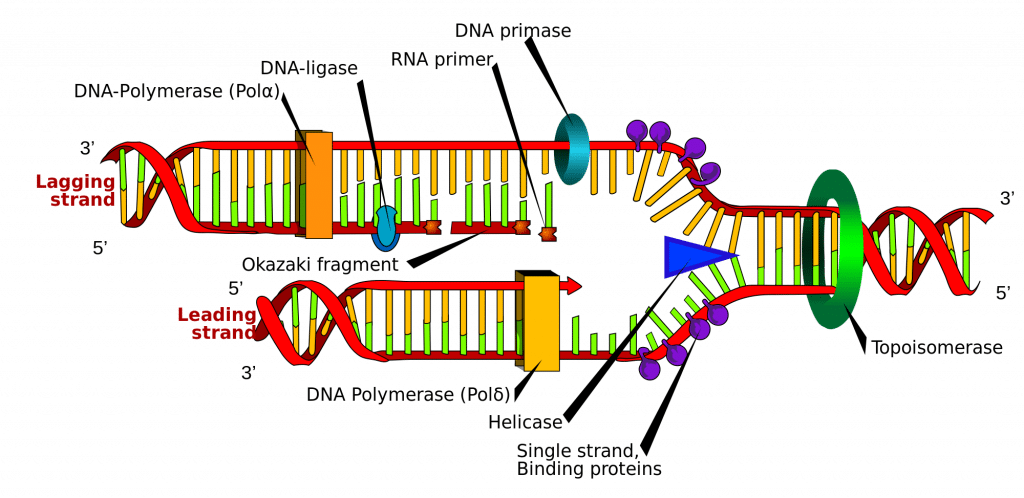 Fig 2.0 – Représentation schématique de la réplication de l’ADN
Fig 2.0 – Représentation schématique de la réplication de l’ADN
Pertinence clinique – Anémie falciforme
L’anémie falciforme est une affection autosomique récessive qui est causée par une substitution d’une seule base, dans laquelle une seule base est changée pour une autre. Dans certains cas, cela peut entraîner une « mutation silencieuse » dans laquelle le gène global n’est pas affecté, mais dans des maladies telles que la drépanocytose, cela se traduit par le brin codant pour une protéine différente.
Dans ce cas, une base adénine est échangée contre une base thymine dans l’un des gènes codant pour l’hémoglobine ; il en résulte que l’acide glutamique est remplacé par la valine. Lorsque ce gène est transcrit en une chaîne polypeptidique, les propriétés qu’il possède sont radicalement modifiées, car l’acide glutamique est hydrophile, tandis que la valine est hydrophobe. Cette région hydrophobe fait que l’hémoglobine a une structure anormale qui peut provoquer des blocages de capillaires conduisant à l’ischémie et potentiellement à la nécrose des tissus et des organes – c’est ce qu’on appelle une crise vaso-occlusive.
Ces crises sont généralement gérées avec une variété de médicaments contre la douleur, y compris des opioïdes et des AINS en fonction de la gravité. Des transfusions de globules rouges peuvent être nécessaires en cas d’urgence, par exemple si le blocage se produit dans les poumons.
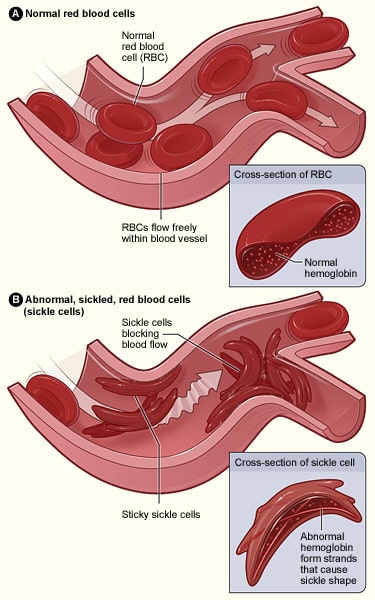 Fig 3.0 – La différence de structure entre les globules rouges normaux, et ceux qui sont affectés par la drépanocytose.
Fig 3.0 – La différence de structure entre les globules rouges normaux, et ceux qui sont affectés par la drépanocytose.