A DNS replikáció, más néven félkonzervatív replikáció, az a folyamat, amelynek során a DNS lényegében megduplázódik. Ez egy fontos folyamat, amely az osztódó sejtben zajlik.
Ebben a cikkben röviden megnézzük a DNS szerkezetét, a DNS replikációjának pontos lépéseit (iniciáció, megnyúlás és termináció), valamint azokat a klinikai következményeket, amelyek akkor jelentkezhetnek, ha ez elromlik.
DNS szerkezete
A DNS több millió nukleotidból áll. Ezek olyan molekulák, amelyek egy dezoxiribóz cukorból állnak, amelyhez egy foszfát és egy bázis (vagy nukleobázis) kapcsolódik. Ezek a nukleotidok foszfodiészterkötésekkel kötődnek egymáshoz szálakba, így alkotva egy “cukor-foszfát gerincet”. A kötés az egyik nukleotid dezoxiribózcukor harmadik szénatomja (a továbbiakban: 3′) és a következő nukleotid egy másik cukor ötödik szénatomja (a továbbiakban: 5′) között jön létre.
N.B.: a 3′ kiejtése “három prím”, az 5′ kiejtése “öt prím”.
A két szál egymással ellentétes vagy antiparallel irányban fut. Ezek a szál teljes hosszában egymáshoz kapcsolódnak az egyes nukleotidok bázisain keresztül. A DNS-hez 4 különböző bázis kapcsolódik; citozin, guanin, adenin és timin. A normál DNS-szálakban a citozin a guaninhoz, az adenin pedig a timinhez kötődik. A két szál együtt kettős spirált alkot.
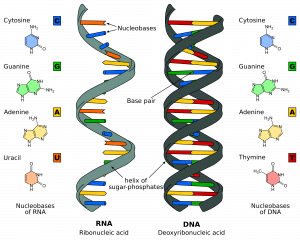 1.0. ábra – Az RNS és a DNS szerkezete
1.0. ábra – Az RNS és a DNS szerkezeteA DNS-replikáció szakaszai
A DNS-replikáció három szakaszban képzelhető el: Kezdet, Nyúlás, Befejezés
Kezdet
A DNS-szintézis a DNS-szál bizonyos pontjain, az úgynevezett “origókon”, azaz meghatározott kódoló régiókban indul. Ezeket az origókat az iniciátorfehérjék célozzák meg, amelyek további fehérjéket toboroznak, amelyek segítik a replikációs folyamatot, és a DNS-eredet körül replikációs komplexet alkotnak. Több origóhely van, és amikor a DNS replikációja megkezdődik, ezeket a helyeket replikációs villáknak nevezik.
A replikációs komplexen belül található a DNS-helikáz enzim, amely feltekeri a kettős spirált, és a két szál mindegyikét feltárja, így azok sablonként használhatók a replikációhoz. Ezt úgy teszi, hogy hidrolizálja a nukleobázisok közötti kötések kialakításához használt ATP-t, így felbontja a két szálat összetartó kötést.
A DNS-primáz egy másik enzim, amely fontos a DNS-replikációban. Kis RNS-primereket szintetizál, amelyek a DNS-polimeráz számára “indítóként” szolgálnak. A DNS-polimeráz az az enzim, amely végső soron az új DNS-szálak létrehozásáért és bővítéséért felelős.
Hosszabbítás
Mihelyt a DNS-polimeráz az eredeti, kibontott két DNS-szálhoz (azaz a sablonszálakhoz) kapcsolódott, képes elkezdeni az új DNS szintetizálását a sablonokhoz igazodva. Lényeges megjegyezni, hogy a DNS-polimeráz csak úgy képes meghosszabbítani a primert, hogy a 3′ végéhez szabad nukleotidokat ad hozzá.
A sablonok egyikét 3′-5′ irányban olvassa be, ami azt jelenti, hogy az új szál 5′-3′ irányban fog kialakulni. Ezt az újonnan képződött szálat nevezzük vezető szálnak. Ezen a szál mentén a DNS-primáznak csak egyszer, az elején kell szintetizálnia egy RNS-primer-t, hogy elindítsa a DNS-polimerázt. Ez azért van így, mert a DNS-polimeráz a sablon 3′-tól 5′-ig történő leolvasásával képes meghosszabbítani az új DNS-szálat, a fent említett 5′-tól 3′-ig irányban szintetizálva.
A másik sablonszál (a lemaradó szál) azonban antiparalel, és ezért 5′-tól 3′-ig irányban olvasható. A folyamatos DNS-szintézisnek, mint a vezető szálnak, 3′ – 5′ irányban kellene történnie, ami lehetetlen, mivel nem tudunk bázisokat hozzáadni az 5′ végéhez. Ehelyett, ahogy a spirál kitekeredik, az RNS-primerek hozzáadódnak a lemaradó szál újonnan feltárt bázisaihoz, és a DNS-szintézis töredékekben történik, de továbbra is az 5′ és 3′ közötti irányban, mint korábban. Ezeket a fragmentumokat Okazaki-fragmentumoknak nevezzük.
Termináció
Az új DNS-szálak bővülésének folyamata addig folytatódik, amíg vagy nem marad több DNS-templát a replikációhoz (azaz a kromoszóma végén), vagy két replikációs villa találkozik, és ezt követően befejeződik. A két replikációs villa találkozása nem szabályozott, és véletlenszerűen történik a kromoszóma folyamán.
Amikor a DNS-szintézis befejeződött, fontos, hogy az újonnan szintetizált szálak megkötődjenek és stabilizálódjanak. A lemaradó szál tekintetében ehhez két enzimre van szükség; az RNAáz H eltávolítja az egyes Okazaki-fragmentumok elején lévő RNS-primereket, a DNS-ligáz pedig összekapcsolja a fragmentumokat, hogy egy teljes szál jöjjön létre.
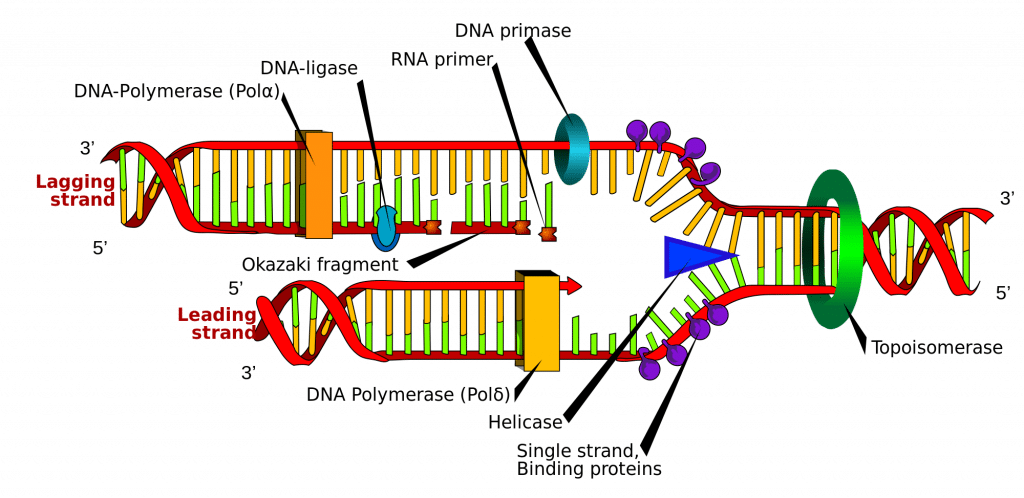 2. ábra.0 – A DNS-replikáció sematikus ábrázolása
2. ábra.0 – A DNS-replikáció sematikus ábrázolása
Klinikai jelentőség – Sarlósejtes vérszegénység
A sarlósejtes vérszegénység egy autoszomális recesszív állapot, amelyet egyetlen bázis cseréje okoz, amelyben csak egy bázis cserélődik ki egy másikra. Egyes esetekben ez egy “csendes mutációt” eredményezhet, amelyben a teljes gén nem érintett, azonban az olyan betegségekben, mint a sarlósejtes vérszegénység, azt eredményezi, hogy a génszál egy másik fehérjét kódol.
Ez esetben a hemoglobint kódoló egyik génben egy adenin bázist egy timin bázisra cserélnek; ez azt eredményezi, hogy a glutaminsav helyébe valin lép. Amikor ez átíródik egy polipeptidlánccá, annak tulajdonságai gyökeresen megváltoznak, mivel a glutaminsav hidrofil, míg a valin hidrofób. Ez a hidrofób régió azt eredményezi, hogy a hemoglobin abnormális szerkezettel rendelkezik, ami a kapillárisok elzáródását okozhatja, ami a szövetek és szervek iszkémiájához és esetleges elhalásához vezethet – ezt nevezik vazo-occlusiv krízisnek.
Ezeket a kríziseket általában különböző fájdalomcsillapítókkal kezelik, beleértve a súlyosságtól függően opioidokat és NSAID-okat. Vörösvérsejt-transzfúzióra szükség lehet vészhelyzetben, például ha az elzáródás a tüdőben jelentkezik.
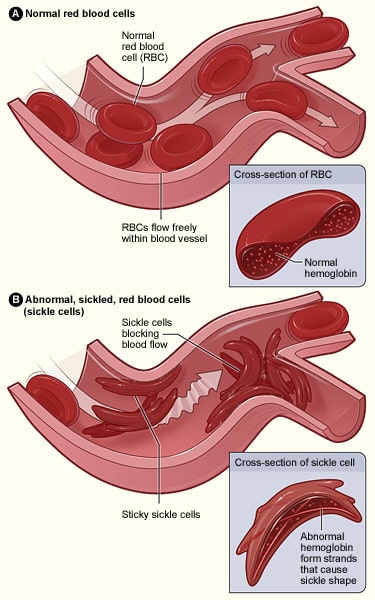 3.0. ábra – A normál vörösvértestek és a sarlósejtes betegségben szenvedők közötti szerkezeti különbség.
3.0. ábra – A normál vörösvértestek és a sarlósejtes betegségben szenvedők közötti szerkezeti különbség.