DNA replicatie, ook bekend als semi-conservatieve replicatie, is het proces waarbij DNA in wezen wordt verdubbeld. Het is een belangrijk proces dat plaatsvindt binnen de delende cel.
In dit artikel zullen we kort ingaan op de structuur van DNA, op de precieze stappen die betrokken zijn bij het repliceren van DNA (initiatie, elongatie en terminatie), en op de klinische gevolgen die kunnen optreden wanneer dit fout gaat.
DNA-structuur
DNA is opgebouwd uit miljoenen nucleotiden. Dit zijn moleculen die zijn opgebouwd uit een deoxyribose suiker, met daaraan vast een fosfaat en een base (of nucleobase). Deze nucleotiden zijn in strengen aan elkaar gehecht via fosfodiesterbindingen om een “suiker-fosfaat ruggengraat” te vormen. De gevormde binding is tussen het derde koolstofatoom op de deoxyribose suiker van een nucleotide (voortaan bekend als de 3′) en het vijfde koolstofatoom van een andere suiker op de volgende nucleotide (bekend als de 5′).
N.B.: 3′ wordt uitgesproken als ‘drie priem’ en 5′ wordt uitgesproken als ‘vijf priem’.
Er zijn twee strengen die in tegengestelde of antiparallelle richtingen ten opzichte van elkaar lopen. Deze zijn over de hele lengte van de streng aan elkaar verbonden door de basen op elke nucleotide. Er zijn 4 verschillende basen verbonden met DNA; Cytosine, Guanine, Adenine, en Thymine. In normale DNA-strengen bindt Cytosine zich aan Guanine, en Adenine aan Thymine. De twee strengen vormen samen een dubbele helix.
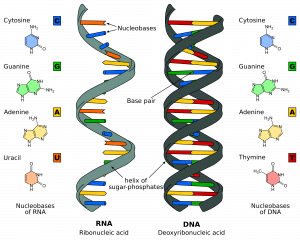 Fig 1.0 – De structuur van RNA en DNA
Fig 1.0 – De structuur van RNA en DNAFasen van DNA-replicatie
DNA-replicatie kan in drie fasen worden opgedeeld; Initiatie, Elongatie, Terminatie
Initiatie
DNA-synthese wordt gestart op bepaalde punten in de DNA-streng die bekend staan als ‘origins’, wat specifieke coderende gebieden zijn. Deze origins zijn het doelwit van initiator-eiwitten, die vervolgens meer eiwitten rekruteren die helpen bij het replicatieproces, en zo een replicatiecomplex vormen rond de DNA-herkomst. Er zijn meerdere oorsprongsgebieden, en wanneer de replicatie van DNA begint, worden deze gebieden replicatievorken genoemd.
In het replicatiecomplex bevindt zich het enzym DNA-helicase, dat de dubbele helix afwikkelt en elk van de twee strengen blootlegt, zodat zij kunnen worden gebruikt als een sjabloon voor replicatie. Het doet dit door het ATP dat gebruikt wordt om de bindingen tussen de nucleobasen te vormen, te hydrolyseren, waardoor de binding die de twee strengen bijeenhoudt, wordt verbroken.
DNA Primase is een ander enzym dat belangrijk is bij de DNA replicatie. Het synthetiseert een kleine RNA primer, die fungeert als een “kick-starter” voor DNA Polymerase. DNA Polymerase is het enzym dat uiteindelijk verantwoordelijk is voor het maken en uitbreiden van de nieuwe strengen DNA.
Uitbreiding
Als het DNA Polymerase zich eenmaal heeft gehecht aan de oorspronkelijke, uitgepakte twee strengen DNA (d.w.z. de sjabloonstrengen), kan het beginnen met de synthese van het nieuwe DNA dat overeenkomt met de sjablonen. Het is van essentieel belang op te merken dat DNA-polymerase de primer alleen kan verlengen door vrije nucleotiden aan het 3′-uiteinde toe te voegen.
Een van de sjablonen wordt in een 3′-naar-5′-richting afgelezen, wat betekent dat de nieuwe streng in een 5′-naar-3′-richting zal worden gevormd. Deze nieuw gevormde streng wordt de leidende streng genoemd. Langs deze streng hoeft DNA Primase maar één keer, aan het begin, een RNA primer te synthetiseren om DNA Polymerase op gang te brengen. Dit komt omdat DNA-polymerase in staat is de nieuwe DNA-streng te verlengen door de template van 3′ naar 5′ te lezen, waarbij de synthese plaatsvindt in een 5′ naar 3′ richting, zoals hierboven is opgemerkt.
De andere template-streng (de achterblijvende streng) is echter antiparallel, en wordt daarom in een 5′ naar 3′ richting gelezen. Continue DNA synthese, zoals in de leidende streng, zou in de 3′ naar 5′ richting moeten plaatsvinden, wat onmogelijk is omdat we geen basen kunnen toevoegen aan het 5′ eind. In plaats daarvan worden tijdens het afwikkelen van de helix RNA-primers toegevoegd aan de nieuw blootgelegde basen op de achterblijvende streng en vindt de DNA-synthese plaats in fragmenten, maar nog steeds in de 5′-3′-richting als voorheen. Deze fragmenten staan bekend als Okazaki-fragmenten.
Terminatie
Het proces van uitbreiding van de nieuwe DNA-strengen gaat door totdat er ofwel geen DNA-sjabloon meer over is om te repliceren (d.w.z. aan het eind van het chromosoom), ofwel twee replicatievorken elkaar ontmoeten en vervolgens worden beëindigd. Het samenkomen van twee replicatievorken is niet gereguleerd en gebeurt willekeurig in de loop van het chromosoom.
Als de DNA-synthese is voltooid, is het belangrijk dat de nieuw gesynthetiseerde strengen worden gebonden en gestabiliseerd. Voor de achterblijvende streng zijn twee enzymen nodig om dit te bereiken; RNAase H verwijdert de RNA-primer die zich aan het begin van elk Okazaki-fragment bevindt, en DNA Ligase voegt de fragmenten samen tot één volledige streng.
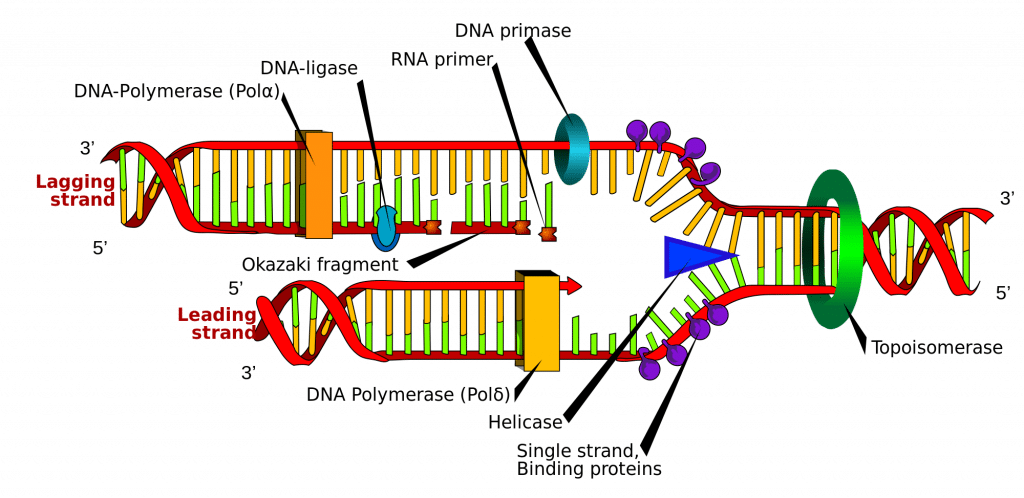 Fig 2.0 – Diagrammatische weergave van DNA-replicatie
Fig 2.0 – Diagrammatische weergave van DNA-replicatie
Clinische relevantie – Sikkelcelanemie
Sikkelcelanemie is een autosomaal recessieve aandoening die wordt veroorzaakt door een enkele basensubstitutie, waarbij slechts één base voor een andere wordt verwisseld. In sommige gevallen kan dit resulteren in een ‘stille mutatie’ waarbij het totale gen niet wordt aangetast, maar bij ziekten zoals sikkelcelanemie resulteert het in de streng die codeert voor een ander eiwit.
In dit geval wordt een adeninebase verwisseld voor een thyminebase in een van de genen die coderen voor hemoglobine; dit resulteert in glutaminezuur dat wordt vervangen door valine. Wanneer dit wordt getranscribeerd in een polypeptideketen, veranderen de eigenschappen ervan radicaal, aangezien glutaminezuur hydrofiel is, terwijl valine hydrofoob is. Dit hydrofobe gebied leidt ertoe dat hemoglobine een abnormale structuur heeft die verstopping van de haarvaten kan veroorzaken met als gevolg ischemie en mogelijk necrose van weefsels en organen – dit staat bekend als een vaso-occlusieve crisis.
Deze crises worden doorgaans behandeld met een verscheidenheid aan pijnmedicatie, waaronder opioïden en NSAID’s, afhankelijk van de ernst. In noodgevallen kunnen transfusies van rode bloedcellen nodig zijn, bijvoorbeeld als de blokkade zich in de longen voordoet.
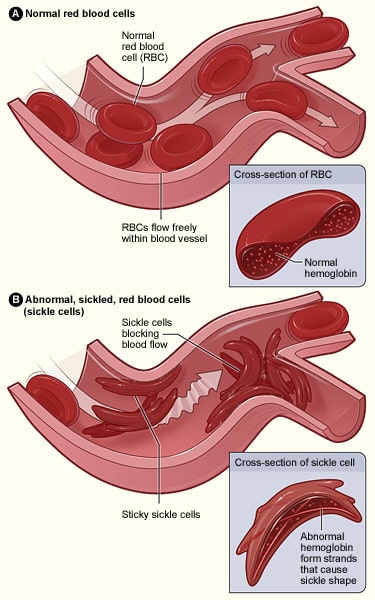 Fig 3.0 – Het verschil in structuur tussen normale rode bloedcellen en cellen die zijn aangetast door sikkelcelziekte.
Fig 3.0 – Het verschil in structuur tussen normale rode bloedcellen en cellen die zijn aangetast door sikkelcelziekte.