Sais biliares e ácidos biliares são derivados polares do colesterol, e representam a principal via para a eliminação do esteróide do corpo.
São moléculas com estruturas semelhantes, mas não idênticas, e características físicas e biológicas diversas.
Sintetizadas no fígado, armazenadas na vesícula biliar, secretadas no duodeno, e finalmente, em sua maioria, reabsorvidas no íleo.
Porque a pH fisiológico estas moléculas estão presentes como ânions, os termos ácido biliar e sais biliares são aqui utilizados como sinônimos.
CONTENTES
- Estrutura química dos sais biliares
- Primário, sais biliares conjugados e secundários
- Função dos ácidos biliares
- Circulação enterohepática dos sais biliares
- >
- Babolimetria intestinal dos ácidos biliares
- Fibras solúveis e reabsorção de sais biliares
>
- Síntese de ácidos biliares primários
- A via clássica ou neutra
- A via alternativa ou ácida
- Vias minoritárias
- Sais biliares: regulação da síntese
>
>
- Estrutura química dos sais biliares
- Sais biliares primários, conjugados e secundários
- Função dos ácidos biliares
- Circulação enterohepática dos sais biliares
- Babolismo intestinal dos ácidos biliares
- Fibras solúveis e reabsorção de sais biliares
- Síntese dos ácidos biliares primários
- O caminho clássico ou neutro
- A via alternativa ou ácida
- Caminhos menores
- Sais biliares: regulação da síntese
Estrutura química dos sais biliares
>
Os sais biliares têm semelhanças e diferenças com a molécula de colesterol.
Como o esteróide, têm um núcleo composto por quatro anéis fundidos: três anéis de cicloexano, rotulados A, B e C, e um anel de cicloopentano, rotulado D. Esta estrutura é o perhydrocyclopentanophenanthrene, mais comumente conhecido como núcleo esteróide.
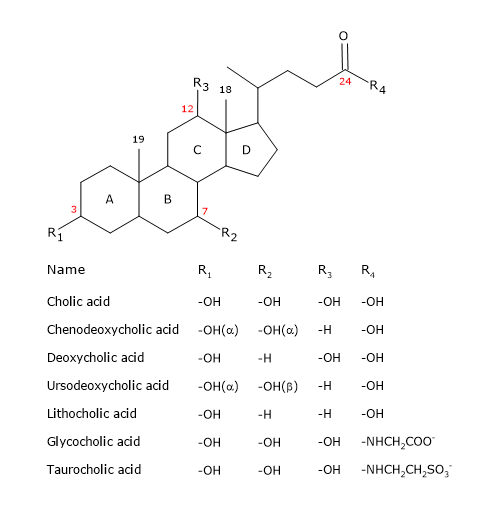
Em vertebrados superiores, eles têm 24 átomos de carbono, já que a cadeia lateral é três carbonos mais curta que a original. Em vertebrados inferiores, os ácidos biliares têm 25, 26, ou 27 átomos de carbono. A cadeia lateral termina com um grupo carboxilo, ionizado a pH 7, que pode ser ligado ao aminoácido glicina ou taurina (ver abaixo).
Além do grupo hidroxila na posição 3, eles têm grupos hidroxila nas posições 7 e/ou 12,
Tudo isso os torna muito mais polares do que o colesterol.
Desde que os anéis A e B são fundidos na configuração cis, a estrutura planar do núcleo esteróide é curva, e é possível identificar:
- um lado côncavo, que é hidrofílico porque os grupos hidroxila e o grupo carboxilo da cadeia lateral, com ou sem o aminoácido ligado, são orientados para ele;
- um lado convexo, que é hidrofóbico porque os grupos metilo presentes na posição 18 e 19 são orientados para ele.
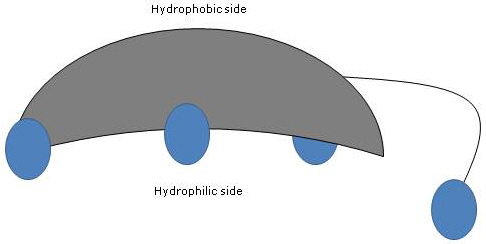
Assim, tendo tanto grupos polares como não polares, são moléculas anfifílicas e excelentes surfactantes. Contudo, a sua estrutura química torna-os diferentes de muitos outros surfactantes, muitas vezes compostos por uma região da cabeça polar e uma cauda não polar.
Sais biliares primários, conjugados e secundários
Ácidos biliares primários são aqueles sintetizados directamente a partir do colesterol nos hepatócitos. Em humanos, os mais importantes são o ácido chólico e o ácido fenodeoxicólico, que constituem 80% de todos os ácidos biliares. Antes de serem secretados na árvore biliar, são quase completamente conjugados, até 98%, com a glicina ou taurina, para formar glicoconjugados e tauroconjugados, respectivamente. Em particular, aproximadamente 75% do ácido chólico e do ácido fenodeoxicólico são conjugados com a glicina, para formar ácido glicocólico e ácido glicochenodeoxicólico, os restantes 25% com a taurina, para formar ácido taurochólico e taurochenodeoxicólico.
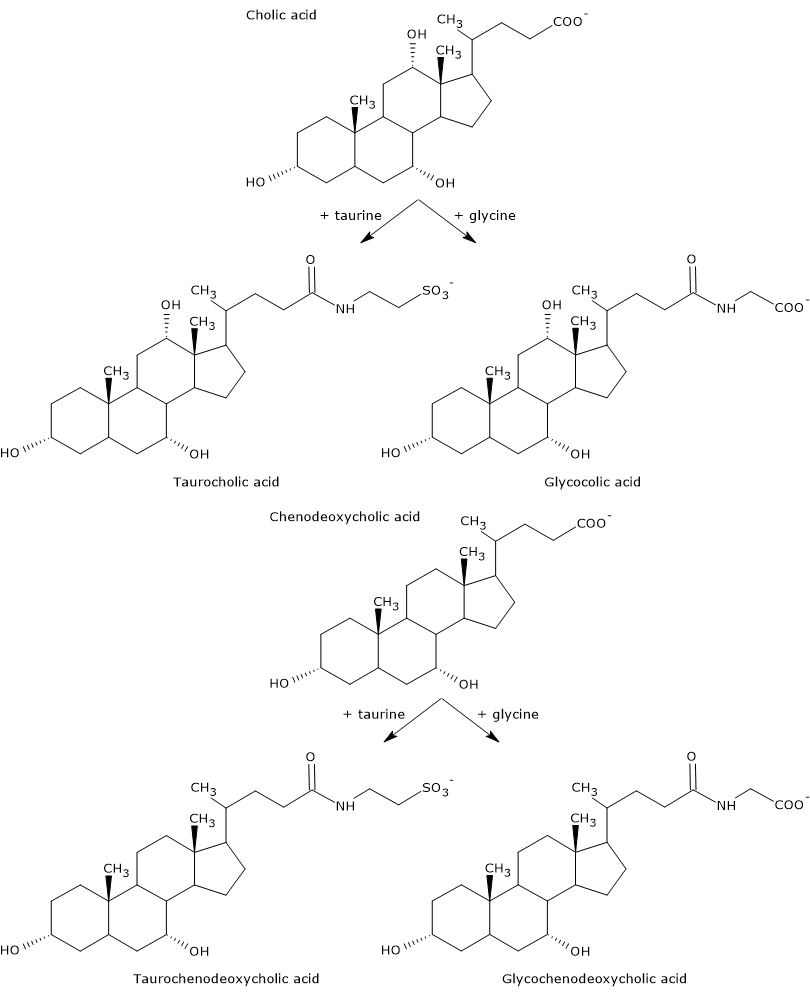
Ácidos biliares conjugados são moléculas com mais grupos hidrofílicos que os ácidos biliares não conjugados, portanto com maior capacidade emulsificante. Na verdade, a conjugação diminui a pKa dos ácidos biliares, de cerca de 6, um valor típico das moléculas não conjugadas, para cerca de 4 para o ácido glicocólico, e cerca de 2 para o ácido taurocólico. Isto faz com que os ácidos biliares conjugados sejam ionizados numa gama mais ampla de pH para formar os sais correspondentes.
A hidrofilicidade do ácido comum e dos sais biliares diminui na seguinte ordem: conjugado com glicina < conjugado com taurina < ácido litocólico < ácido deoxicólico < ácido fenodeoxicólico < ácido fenodeoxicólico < ácido chicólico < ácido ursodeoxicólico.
Finalmente, a conjugação também diminui a citotoxicidade dos ácidos biliares primários.
Ácidos biliares secundários são formados a partir de ácidos biliares primários que não foram reabsorvidos a partir do intestino delgado. Quando chegam ao cólon, podem sofrer várias modificações por microbiota intestinal para formar ácidos biliares secundários (ver abaixo). Eles compõem os restantes 20% do pool de ácidos biliares do organismo.
Uma outra forma de classificar os sais biliares é baseada na sua conjugação com glicina e taurina e no seu grau de hidroxilação. Nesta base, três categorias são identificadas.
- Conjugados trihidroxi, como ácido taurochólico e ácido glicocólico.
- Conjugados dihidroxi, como ácido glicodeoxicólico, ácido glicochenodeoxicólico, ácido taurochenodeoxicólico, e ácido taurodeoxicólico. Eles representam cerca de 60% dos sais biliares presentes na bílis.
- Formas não conjugadas, como ácido cholic, ácido deoxicólico, ácido fenodeoxicólico, e ácido litocólico.
Função dos ácidos biliares
Todas as suas funções fisiológicas são realizadas na forma conjugada.
- São a principal via para a eliminação do colesterol do corpo humano.
Indeed, os humanos não têm as enzimas para quebrar os anéis de ciclohexano ou o anel ciclopentano do núcleo do esteróide, nem para oxidar o colesterol ao CO2 e à água.
O outro mecanismo para eliminar o esteróide do corpo é como o colesterol per se na bílis. - Os sais biliares são surfactantes fortes. E em particular, os conjugados di- e tri-hidroxi são os melhores surfactantes entre os ácidos biliares, muito mais eficazes que os não conjugados, uma vez que têm mais grupos polares.
Em contacto com os lípidos apolares na luz do intestino delgado, a superfície apolar convexa interage com os lípidos apolares, tais como triglicéridos, ésteres de colesterol e ésteres de vitaminas lipossolúveis, enquanto que a superfície polar côncava interage com o meio aquoso circundante. Isto aumenta a dispersão dos lípidos apolares no meio aquoso, pois permite a formação de pequenas gotículas lipídicas, aumentando a área superficial para:
atividade da lipase, principalmente a lipase pancreática, (os sais biliares também desempenham um papel direto na ativação desta enzima);
atividade da esterase intestinal.
Subsequentemente, facilitam a absorção de produtos de digestão lipídica, bem como de vitaminas lipossolúveis pela mucosa intestinal graças à formação de micelas mistas.
Os ácidos biliares desempenham uma função semelhante na vesícula biliar onde, formando micelas mistas com fosfolípidos, evitam a precipitação do colesterol.
Nota: Como consequência da disposição dos grupos polares e não polares, os ácidos biliares formam micelas em solução aquosa, geralmente composta de menos de 10 monómeros, desde que a sua concentração esteja acima da chamada concentração micelar crítica ou CMC.
- A nível intestinal, eles modulam a secreção das enzimas pancreáticas e colecistoquinina.
- No intestino delgado e grosso, eles têm uma potente atividade antimicrobiana, principalmente ácido deoxicólico, em particular contra bactérias Gram-positivas. Esta actividade pode ser devida a danos no DNA oxidativo, e/ou a danos na membrana celular. Portanto, elas desempenham um papel importante na prevenção do crescimento excessivo de bactérias, mas também na regulação da composição da microbiota intestinal.
- Nos últimos anos, torna-se evidente o seu papel regulador no controlo do metabolismo energético, e em particular no manuseamento da glucose hepática.
Circulação enterohepática dos sais biliares
Após a ingestão de gordura, as células enteroendócrinas do duodeno secretam a colecistequinina na corrente sanguínea. A ligação hormonal aos receptores das células musculares lisas da vesícula biliar promove sua contração; o hormônio também causa o relaxamento do esfíncter de Oddi. Tudo isto resulta na secreção da bílis, e portanto dos ácidos biliares para o duodeno.
Em condições fisiológicas, o pool de sal da bílis humana é constante, e igual a cerca de 3-5 g. Isto é possível por dois processos:
- a sua reabsorção intestinal;
- a sua síntese de novo (ver abaixo).
Sais biliares segregados a 95% são reabsorvidos do intestino, não juntamente com os produtos da digestão lipídica, mas através de um processo chamado circulação enterohepática.
É um sistema de reciclagem extremamente eficiente, que parece ocorrer pelo menos duas vezes para cada refeição, e inclui o fígado, a árvore biliar, o intestino delgado, o cólon, e a circulação portal através da qual as moléculas reabsorvidas retornam ao fígado. Tal recirculação é necessária uma vez que a capacidade do fígado para sintetizar os ácidos biliares é limitada e insuficiente para satisfazer as necessidades intestinais se os sais biliares fossem excretados nas fezes em grandes quantidades.
A maior parte dos sais biliares são reabsorvidos no íleo distal, a parte inferior do intestino delgado, por um transportador dependente de sódio dentro da borda da escova dos enterócitos, chamado transportador de ácido biliar dependente de sódio ou ASBT, que realiza o co transporte de uma molécula de ácido biliar e dois íons de sódio.
Com o enterócito, pensa-se que os ácidos biliares são transportados através do citosol até à membrana basolateral pela proteína ligadora de ácido biliar ileal ou IBABP. Eles atravessam a membrana basolateral pelo transportador de soluto orgânico alfa-beta ou OSTβ/OSTβ, passam para a circulação portal e, ligados à albumina, atingem o fígado.
Deve-se notar que uma pequena porcentagem de ácidos biliares atinge o fígado através da artéria hepática.
Um nível hepático, sua extração é muito eficiente, com uma fração de extração de primeira passagem variando de 50 a 90%, uma porcentagem que depende da estrutura ácida da bílis. A captação de ácidos biliares conjugados é mediada principalmente por um sistema de transporte ativo dependente de Na+, ou seja, o polipéptido cotransportador de taurocolato dependente de sódio ou NTCP. Entretanto, também pode ocorrer uma absorção independente de sódio, realizada por proteínas da família dos polipeptídeos transportadores de ânions orgânicos ou OATP, principalmente OATP1B1 e OATP1B3.
A etapa limitadora da taxa na circulação enterohepática é sua secreção canalicular, em grande parte mediada pela bomba de exportação de sal biliar ou BSEP, em um processo dependente de ATP. Esta bomba transporta sais biliares monoanionicos, que são os mais abundantes. Os ácidos biliares conjugados com ácido glucurônico ou sulfato, que são dianionicos, são transportados por diferentes portadores, como MRP2 e BCRP.
Nota: Os níveis séricos de ácidos biliares variam em função da taxa de reabsorção dos mesmos e, portanto, são mais altos durante as refeições, quando a circulação enterohepática é mais ativa.
Babolismo intestinal dos ácidos biliares
Ácidos biliares que escapam da absorção ileal passam para o cólon onde são parcialmente modificados pela microbiota intestinal e são convertidos em ácidos biliares secundários.
As principais reacções são listadas abaixo.
- Desconjugação
Na cadeia lateral, pode ocorrer hidrólise da ligação C24 N-acyl amide, com libertação de ácidos biliares não conjugados e glicina ou taurina. Esta reação é catalisada por hidrolases bacterianas presentes tanto no intestino delgado quanto no cólon. - 7α-Dehidroxilation
Quantitativamente, é a reação mais importante, realizada por desidratases bacterianas colônicas que removem o grupo hidroxila na posição 7 para formar ácidos biliares 7-deoxídicos. Em particular, o ácido deoxicólico é formado a partir do ácido chólico, e o ácido litocólico, um ácido biliar secundário tóxico, a partir do ácido fenodeoxicólico.
Deve-se notar que 7α-dehydroxylation, ao contrário da oxidação e epimerização (ver abaixo), só pode ocorrer em ácidos biliares não conjugados, e portanto, a desconjugação é um pré-requisito essencial. - Oxidação e epimerização
São reacções envolvendo os grupos hidroxila nas posições 3, 7 e 12, catalisadas por desidrogenase hidroxisteróide bacteriana. Por exemplo, o ácido ursodeoxicólico deriva da epimerização do ácido fenodeoxicólico.
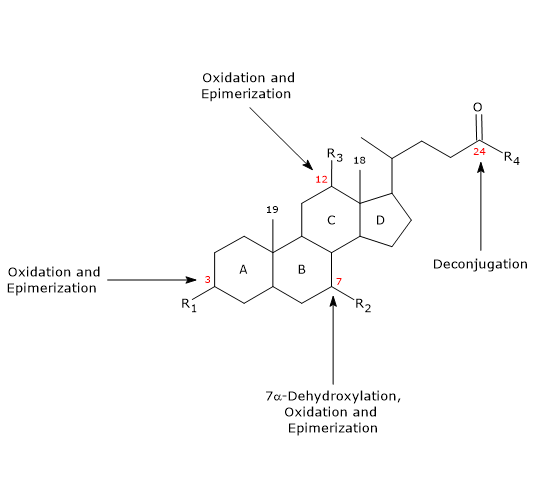
Alguns dos ácidos biliares secundários são então reabsorvidos do cólon e retornam ao fígado. Nos hepatócitos, eles são conjugados novamente, se necessário, e ressecados. Aqueles que não são reabsorvidos, são excretados nas fezes.
Quando as oxidações e desconjugações são realizadas por um amplo espectro de bactérias anaeróbias, 7α-des-hidroxiolações são realizadas por um número limitado de anaeróbios do cólon.
7α>As oxidações e desconjugações aumentam a pKa dos ácidos biliares, e portanto a sua hidrofobicidade, permitindo um certo grau de absorção passiva através da parede do cólon.
O aumento da hidrofobicidade está também associado a um aumento da toxicidade destas moléculas. E uma alta concentração de ácidos biliares secundários na bílis, sangue e fezes tem sido associada à patogênese do câncer de cólon.
Fibras solúveis e reabsorção de sais biliares
A reabsorção de sais biliares pode ser reduzida pela ação quelante das fibras solúveis, como as encontradas em frutas frescas, legumes, aveia e farelo de aveia, que as ligam, diminuindo sua absorção. Por sua vez, isso aumenta a síntese do ácido biliar de novo, aumentando a expressão do 7α-hidroxilase e esterol 12α-hidroxilase (veja abaixo), e assim reduz a concentração de colesterol hepatócito.
O esgotamento do colesterol hepático aumenta a expressão do receptor LDL, e assim reduz a concentração plasmática do colesterol LDL. Por outro lado, também estimula a síntese de HMG-CoA redutase, a enzima chave na biossíntese do colesterol.
Nota: Alguns medicamentos anti-colesterol atuam ligando os ácidos biliares no intestino, impedindo sua reabsorção.
Síntese dos ácidos biliares primários
Quantitativamente, os ácidos biliares são o principal produto do metabolismo do colesterol.
Como dito anteriormente, a circulação enterohepática e sua síntese de novo mantém um tamanho constante do pool de ácidos biliares. Em particular, a síntese de novo permite a substituição dos sais biliares excretados na face, cerca de 5-10% do pool corporal, ou seja ~ 0,5 g/dia.
Below, a síntese de ácido chólico e ácido fenodeoxicólico, e sua conjugação com os aminoácidos taurina e glicina, é descrita.
Existem duas vias principais para a síntese de ácido biliar: a via clássica e a via alternativa. Além disso, alguns outros caminhos menores também serão descritos.
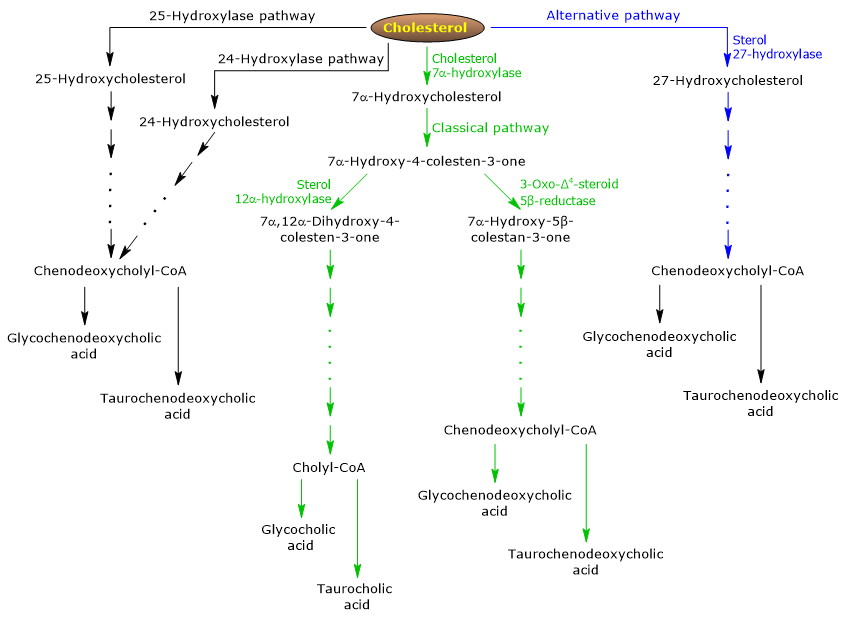
O caminho clássico ou neutro
Em humanos, até 90% dos sais biliares são produzidos através do caminho clássico (ver fig. 5), também referido como caminho “neutro” uma vez que os intermediários são moléculas neutras.
É uma via metabólica presente apenas no fígado, que consiste de reações catalisadas por enzimas localizadas no citosol, retículo endoplasmático, peroxissomas e mitocôndrias, e cujos produtos finais são os conjugados de ácido chólico e ácido fenodeoxicólico.
- A primeira reação é a hidroxilação na posição 7 do colesterol, para formar 7α-hidroxicolesterol. A reação é catalisada pelo colesterol 7α-hidroxilase ou CYP7A1 (E.C. 1.14.14.23). É uma enzima localizada no retículo endoplasmático, e catalisa a etapa limitadora da taxa do caminho.
Colesterol + NADPH + H+ + O2 → 7α-Hidroxicolesterol + NADP+ + H2O
- 7α-Hidroxicolesterol sofre a oxidação do grupo 3β-hydroxyl e a mudança da dupla ligação da posição 5,6 para a posição 4,5, para formar 7α-hydroxy-4-cholesten-3-one. A reação é catalisada pela 3β-hydroxy-Δ5-C27-steroid oxidoreductase ou HSD3B7 (E.C. 1.1.1.181), uma enzima localizada no retículo endoplasmático.
- 7α-Hidroxi-4-cholesten-3-one pode seguir duas vias:
entrar no caminho que leva à síntese do ácido chólico, através da reacção catalisada por 7α-hydroxy-4-cholesten-3-one 12α-monooxygenase ou sterol 12α-hydroxylase ou CYP8B1 (E.C. 1.14.18.8), uma enzima localizada no retículo endoplasmático;
para entrar no caminho que conduz à síntese do ácido fenodeoxicólico, através da reação catalisada pelo esteróide 3-oxo-Δ4 5β-reductase ou AKR1D1 (E.C. 1.3.1.3), uma enzima citosólica.
De salientar que a actividade do esterol 12α-hidroxilase determina a razão entre o ácido cólico e o ácido fenodeoxicólico e, em última análise, a capacidade detergente da piscina de ácido biliar. E, de facto, a regulação da transcrição do gene do esterol 12α-hidroxilase é uma das principais etapas reguladoras da via clássica.
Por isso, se 7α-hydroxy-4-cholesten-3-one prosseguir através da reacção catalisada pelo esterol 12α-hidroxilase, as seguintes reacções ocorrerão.
- 7α-Hidroxi-4-colesten-3-ona é hidroxilada na posição 12 por esterol 12α-hidroxilase, para formar 7α,12α-dihydroxy-4-cholesten-3-ona.
- 7α,12α-dihidroxi-4-colesten-3-ona sofre redução da dupla ligação na posição 4,5, na reação catalisada pelo esteróide 3-oxo-Δ4 5β-reductase, para formar 5β-cholestan-7α,12α-diol-3-ona.
- 5β-Cholestan-7α,12α-diol-3-one sofre redução do grupo hidroxila na posição 4, na reação catalisada por 3α-hidroxiesteroida-desidrogenase ou AKR1C4 (EC 1.1.1.213), uma enzima citosólica, para formar 5β-cholestan-3α,7α,12α-triol.
- 5β-Cholestan-3α,7α,12α-triol sofre oxidação da cadeia lateral através de três reacções catalisadas por esterol 27-hidroxilase ou CYP27A1 (EC 1.14.15.15). Trata-se de uma enzima mitocondrial também presente nos tecidos extra-hepáticos e macrófagos, que introduz um grupo hidroxila na posição 27. O grupo hidroxila é oxidado ao aldeído, e depois ao ácido carboxílico, para formar 3α,7α,12α-tri-hidroxi-5β-ácido colestanóico.
- 3α,7α,12α-trihidroxi-5β-ácido colestanóico é ativado à sua coenzima A ester, 3α,7α,12α-trihidroxi-5β-colestanóico, na reação catalisada pela cadeia muito longa acil-CoA sintetase ou VLCS (EC 6.2.1.-), ou ácido biliar CoA sintetase ou BACS (EC 6.2.1.7), ambos localizados no retículo endoplasmático.
- 3α,7α,12α-Trihidroxi-5β-colestanoyl-CoA é transportado para peroxisomas onde sofre cinco reações sucessivas, cada uma catalisada por uma enzima diferente. Nas duas últimas reações, a cadeia lateral é encurtada para quatro átomos de carbono, e finalmente a colilcoA é formada.
- Na última etapa, ocorre a conjugação, via amide bond, do grupo ácido carboxílico da cadeia lateral com o aminoácido glicina ou taurina. A reação é catalisada pelo ácido biliar-CoA:aminoácido N-aciltransferase ou o BAAT (EC 2.3.1.65), que é predominantemente localizado em peroxissomas.
Os produtos da reação são, portanto, os ácidos biliares conjugados: ácido glicocólico e ácido taurocólico.
Se 7α-hydroxy-4-cholesten-3-one não prosseguir através da reação catalisada pelo esterol 12α-hidroxilase, ele entra no caminho que leva à síntese de conjugados de ácido fenodeoxicólico, através das reações descritas abaixo.
- 7α-Hidroxi-4-cholesten-3-one é convertido para 7α-hydroxy-5β-cholestan-3-one na reação catalisada pelo 3-oxo-Δ4-esteróide 5β-reductase.
- 7α-Hidroxi-5β-cholestan-3-one é convertido para 5β-cholestan-3α,7α-diol na reação catalisada pelo 3α-hidroxi-esteróide desidrogenase.
Então, os ácidos biliares conjugados ácido glicochenodeoxicólico e ácido taurochenodeoxicólico são formados por modificações similares àquelas observadas para a conjugação do ácido cholic, e catalisados em sua maioria pelas mesmas enzimas.
Nota: Os ácidos biliares não conjugados formados no intestino devem alcançar o fígado para serem conjugados.
A via alternativa ou ácida
Prevalora no feto e neonato, enquanto que nos adultos leva à síntese de menos de 10% dos sais biliares.
Esta via (ver fig. 5) difere da via clássica em que:
- os produtos intermediários são moléculas ácidas, das quais o nome alternativo “via ácida”;
- a oxidação da cadeia lateral é seguida por modificações do núcleo esteróide, e não vice-versa;
- os produtos finais são conjugados de ácido fenodeoxicólico.
O primeiro passo envolve a conversão do colesterol em 27-hidroxicolesterol na reação catalisada pelo esterol 27-hidroxilase.
27-Hidroxicolesterol pode seguir dois caminhos.
Rota A
- 27-hidroxicolesterol é convertido em 3β-hydroxy-5-ácido colestenóico numa reacção catalisada por esterol 27-hidroxilase.
- 3β-Hidroxi-5-ácido colestenóico é hidroxilado na posição 7 na reação catalisada por oxistrole 7α-hidroxilase ou CYP7B1 (EC 1.14.13.100), uma enzima localizada no retículo endoplasmático, para formar 3β-7α-dihidroxi-5-ácido colestenóico.
- 3β-7α- ácido dihidroxi-5-colestenóico é convertido em ácido 3-oxo-7α-hidroxi-4-colestenóico, na reação catalisada por 3β-hydroxy-Δ5-C27-steroidoreductase.
- 3-Oxo-7α-hidroxi-4-ácido colestenóico, como resultado de modificações da cadeia lateral, forma ácido fenodeoxicólico, e depois seus conjugados.
Rota B
- 27-Hidroxicolesterol é convertido para 7α,27-dihidroxicolesterol na reacção catalisada pelo oxisterol 7α-hidroxilase e colesterol 7α-hidroxilase.
- 7α,27-Diidroxicolesterol é convertido para 7α,26-dihidroxi-4-colesten-3-ona na reação catalisada por 3β-hydroxy-Δ5-C27-steroidoreductase;
7α, 26-Dihidroxi-4-colesten-3-one pode ser transformado diretamente em conjugados de ácido fenodeoxicólico, ou pode ser convertido em ácido 3-oxo-7α-hidroxi-4-colestenóico, e então sofrer modificações na cadeia lateral e outras reações que levam à síntese dos conjugados de ácido fenodeoxicólico.
Caminhos menores
Também existem caminhos menores (veja fig. 5) que contribuem para a síntese do sal biliar, embora em menor extensão que os caminhos clássicos e alternativos.
Por exemplo:
- Um colesterol 25-hidroxilase (EC 1.14.99.38) é expresso no fígado.
- Um colesterol 24-hidroxilase ou CYP46A1 (EC 1.14.14.25) é expresso no cérebro, e portanto, embora o órgão não possa exportar colesterol, ele exporta oxisteróis.
- A não específica 7α-hidroxilase também foi descoberta. Ela é expressa em todos os tecidos e parece estar envolvida na geração de oxisteróis, que podem ser transportados para hepatócitos para serem convertidos em ácido fenodeoxicólico.
Adicionalmente, o esterol 27-hidroxilase é expresso em vários tecidos, e portanto seus produtos de reação devem ser transportados para o fígado para serem convertidos em sais biliares.
Sais biliares: regulação da síntese
Regulação da síntese dos ácidos biliares ocorre através de um mecanismo de feedback negativo, particularmente na expressão do colesterol 7α-hidroxilase e esterol 12α-hidroxilase.
Quando ocorre um excesso de ácidos biliares, tanto livres como conjugados, estas moléculas ligam-se ao receptor nuclear farnesóide X ou FRX, activando-o: o ácido biliar mais eficaz é o ácido fenodeoxicólico, enquanto outras, como o ácido ursodeoxicólico, não o activam.
FRX induz a expressão do parceiro de repressor transcripcional pequeno heterodímero ou SHP, que por sua vez interage com outros factores de transcrição, tais como o receptor hepático homolog-1 ou LRH-1, e o factor nuclear hepatocitário-4α ou HNF-4α. Estes factores de transcrição ligam-se a uma sequência na região promotora dos genes 7α-hydroxylase e 12α-hydroxylase, região chamada elementos de resposta ácida biliar ou BAREs, inibindo a sua transcrição.
Uma das razões pelas quais a síntese de sais biliares é fortemente regulada é porque muitos dos seus metabolitos são tóxicos.
Chiang J.Y.L. Ácidos biliares: regulação da síntese. J Lipid Res 2009;50(10):1955-66. doi:10.1194/jlr.R900010-JLR200
Gropper S.S., Smith J.L. Nutrição avançada e metabolismo humano. Edição 6h. Cengage Learning, 2012
Moghimipour E., Ameri A., e Handali S. Efeitos de aumento da absorção de sais biliares. Molecules 2015;20(8); 14451-73. doi:10.3390/molecules200814451
Monte M.J., Marin J.J.G., Antelo A., Vazquez-Tato J. Ácidos biliares: Química, fisiologia e fisiopatologia. Mundo J Gastroenterol 2009;15(7):804-16. doi:10.3748/wjg.15.804
Rawn J.D. Biochimica. Mc Graw-Hill, Neil Patterson Publishers, 1990
Rosenthal M.D., Glew R.H. Medical biochemistry – Human metabolism in health and disease. John Wiley J. & Sons, Inc., Publication, 2009
Sundaram S.S., Bove K.E., Lovell M.A. e Sokol R.J. Mechanisms of Disease: inborn errors of bile acid synthesis. Nat Clin Pract Gastroenterol Hepatol 2008;5(8):456-68. doi:10.1038/ncpgasthep1179