Geny i chromosomy
Klasyczne zasady genetyki zostały wydedukowane przez Gregora Mendla w 1865 roku, na podstawie wyników eksperymentów hodowlanych z grochem. Mendel badał dziedziczenie wielu dobrze zdefiniowanych cech, takich jak kolor nasion, i był w stanie wydedukować ogólne zasady ich przekazywania. We wszystkich przypadkach mógł poprawnie zinterpretować zaobserwowane wzorce dziedziczenia, zakładając, że każda cecha jest determinowana przez parę dziedziczonych czynników, które obecnie nazywamy genami. Po każdym z rodziców dziedziczona jest jedna kopia genu (zwana allelem) określająca każdą cechę. Na przykład hodowla dwóch odmian grochu – jednej o nasionach żółtych, a drugiej o nasionach zielonych – daje następujące wyniki (rysunek 3.1). Każda z odmian rodzicielskich ma dwie identyczne kopie genu określającego odpowiednio żółte (Y) lub zielone (y) nasiona. Rośliny potomne są zatem hybrydami, które odziedziczyły jeden gen dla nasion żółtych (Y) i jeden dla nasion zielonych (y). Wszystkie te rośliny potomne (pierwsze pokolenie F1) mają żółte nasiona, więc żółte (Y) jest dominujące, a zielone (y) recesywne. Genotyp (skład genetyczny) grochu F1 jest więc Yy, a jego fenotyp (wygląd fizyczny) jest żółty. Jeśli jedno potomstwo F1 jest hodowane z drugim, dając początek potomstwu F2, geny dla żółtych i zielonych nasion segregują w charakterystyczny sposób, tak że stosunek roślin F2 z żółtymi nasionami do tych z zielonymi nasionami wynosi 3:1.
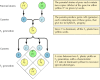
Rysunek 3.1
Dziedziczenie genów dominujących i recesywnych.
Odkrycia Mendla, najwyraźniej wyprzedzające swój czas, były w dużej mierze ignorowane aż do roku 1900, kiedy to prawa Mendla zostały ponownie odkryte, a ich znaczenie uznane. Wkrótce potem zaproponowano rolę chromosomów jako nośników genów. Uświadomiono sobie, że większość komórek roślin wyższych i zwierząt jest diploidalna – zawiera dwie kopie każdego chromosomu. Tworzenie się komórek rozrodczych (plemnika i jaja) wymaga jednak unikalnego rodzaju podziału komórkowego (mejozy), w którym tylko jeden członek każdej pary chromosomów jest przekazywany do każdej komórki potomnej (rysunek 3.2). W konsekwencji plemnik i komórka jajowa są haploidalne, zawierają tylko po jednej kopii każdego chromosomu. Połączenie tych dwóch haploidalnych komórek podczas zapłodnienia tworzy nowy diploidalny organizm, zawierający teraz po jednym członku każdej pary chromosomów pochodzących od męskiego i żeńskiego rodzica. Zachowanie par chromosomów jest więc analogiczne do zachowania genów, co prowadzi do wniosku, że geny są przenoszone na chromosomach.
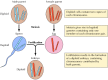
Rysunek 3.2
Chromosomy podczas mejozy i zapłodnienia. Zilustrowano dwie pary chromosomów hipotetycznego organizmu.
Podstawy mutacji, powiązań genetycznych oraz związków między genami i chromosomami zostały w dużej mierze ustalone przez eksperymenty przeprowadzone z muszką owocową, Drosophila melanogaster. Drosophila mogą być łatwo utrzymywane w laboratorium, a oni rozmnażają się około co dwa tygodnie, co jest znaczną zaletą dla eksperymentów genetycznych. Rzeczywiście, te cechy nadal sprawiają, że Drosophila jest organizmem z wyboru dla badań genetycznych zwierząt, zwłaszcza genetycznej analizy rozwoju i różnicowania.
W początkach 1900 roku zidentyfikowano szereg zmian genetycznych (mutacji) w Drosophila, zwykle wpływających na łatwo obserwowalne cechy, takie jak kolor oczu lub kształt skrzydeł. Eksperymenty hodowlane wykazały, że niektóre z genów rządzących tymi cechami dziedziczą się niezależnie od siebie, co sugeruje, że geny te znajdują się na różnych chromosomach, które segregują się niezależnie podczas mejozy (rysunek 3.3). Inne geny są jednak często dziedziczone razem, jako cechy sparowane. Mówi się, że takie geny są ze sobą sprzężone dzięki temu, że znajdują się na tym samym chromosomie. Liczba grup powiązanych genów jest taka sama jak liczba chromosomów (cztery u Drosophila), co potwierdza tezę, że chromosomy są nośnikami genów.

Rysunek 3.3
Segregacja genów i powiązania. (A) Segregacja dwóch hipotetycznych genów dla kształtu (A/a = kwadratowy/okrągły) i koloru (B/b = czerwony/niebieski) zlokalizowanych na różnych chromosomach. (B) Powiązanie dwóch genów zlokalizowanych na tym samym chromosomie.
Połączenie między genami nie jest jednak całkowite; chromosomy wymieniają się materiałem podczas mejozy, co prowadzi do rekombinacji między połączonymi genami (rysunek 3.4). Częstość rekombinacji między dwoma połączonymi genami zależy od odległości między nimi na chromosomie; geny, które są blisko siebie, rekombinują rzadziej niż geny bardziej od siebie oddalone. Częstotliwość rekombinacji różnych genów może być zatem wykorzystana do określenia ich względnego położenia na chromosomie, co umożliwia konstruowanie map genetycznych (rysunek 3.5). Do 1915 roku zdefiniowano i zmapowano prawie sto genów na czterech chromosomach Drosophili, co doprowadziło do ogólnej akceptacji chromosomowej podstawy dziedziczności.
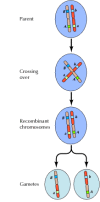
Rysunek 3.4
Rekombinacja genetyczna. Podczas mejozy członkowie par chromosomów wymieniają się materiałem. Wynikiem tego jest rekombinacja między połączonymi genami.
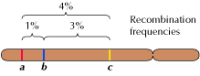
Rysunek 3.5
Mapa genetyczna. Trzy geny są zlokalizowane na hipotetycznym chromosomie w oparciu o częstości rekombinacji między nimi (1% rekombinacji między a i b; 3% między b i c; 4% między a i c). Częstości rekombinacji są w przybliżeniu proporcjonalne (więcej…)