Genes e Cromossomas
Os princípios clássicos da genética foram deduzidos por Gregor Mendel em 1865, com base nos resultados das experiências de reprodução com ervilhas. Mendel estudou a herança de uma série de traços bem definidos, como a cor da semente, e foi capaz de deduzir regras gerais para a sua transmissão. Em todos os casos, ele pôde interpretar corretamente os padrões de herança observados, assumindo que cada traço é determinado por um par de fatores herdados, que agora são chamados de genes. Uma cópia do gene (chamada alelo) especificando cada traço é herdado de cada um dos pais. Por exemplo, reproduzir duas linhagens de ervilhas – uma com sementes amarelas, e a outra verde – produz os seguintes resultados (Figura 3.1). As linhagens parentais têm, cada uma, duas cópias idênticas do gene especificando sementes amarelas (Y) ou verdes (y), respectivamente. As plantas progenitoras são portanto híbridas, tendo herdado um gene para as sementes amarelas (Y) e outro para as sementes verdes (y). Todas estas plantas descendentes (a primeira filial, ou geração F1) têm sementes amarelas, por isso diz-se que o amarelo (Y) é dominante e o verde (y) é recessivo. O genótipo (composição genética) das ervilhas F1 é assim Yy, e o seu fenótipo (aparência física) é amarelo. Se uma descendência F1 é criada com outra, dando origem à descendência F2, os genes das sementes amarelas e verdes segregam de forma característica de tal forma que a proporção entre plantas F2 com sementes amarelas e aquelas com sementes verdes é de 3:1.
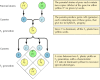
Figure 3.1
Inheritância dos genes dominantes e recessivos.
Os achados de Mendel, aparentemente à frente de seu tempo, foram largamente ignorados até 1900, quando as leis de Mendel foram redescobertas e sua importância reconhecida. Pouco tempo depois, o papel dos cromossomas como portadores de genes foi proposto. Percebeu-se que a maioria das células de plantas e animais superiores são diplóides – contendo duas cópias de cada cromossoma. A formação das células germinativas (o esperma e o óvulo), entretanto, envolve um tipo único de divisão celular (meiose) na qual apenas um membro de cada par cromossômico é transmitido para cada célula progenitora (Figura 3.2). Consequentemente, os espermatozóides e o óvulo são haplóides, contendo apenas uma cópia de cada cromossoma. A união destas duas células haplóides na fertilização cria um novo organismo diplóide, contendo agora um membro de cada par de cromossomas derivado do macho e um do progenitor feminino. O comportamento dos pares de cromossomos é, portanto, paralelo ao dos genes, levando à conclusão de que os genes são carregados nos cromossomos.
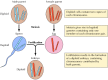
Figure 3.2
Cromossomos na meiose e fertilização. Dois pares de cromossomas de um organismo hipotético são ilustrados.
Os fundamentos da mutação, ligação genética e as relações entre genes e cromossomas foram em grande parte estabelecidos por experimentos realizados com a mosca da fruta, Drosophila melanogaster. A Drosophila pode ser facilmente mantida em laboratório, e reproduzem-se a cada duas semanas, o que é uma vantagem considerável para os experimentos genéticos. De facto, estas características continuam a fazer da Drosophila um organismo de escolha para estudos genéticos de animais, particularmente a análise genética do desenvolvimento e diferenciação.
No início do século XIX, foram identificadas várias alterações genéticas (mutações) na Drosophila, geralmente afectando características facilmente observáveis como a cor dos olhos ou a forma das asas. Experimentos de reprodução indicaram que alguns dos genes que regem esses traços são herdados independentemente uns dos outros, sugerindo que esses genes estão localizados em cromossomos diferentes que se segregam independentemente durante a meiose (Figura 3.3). Outros genes, entretanto, são frequentemente herdados juntos como características pareadas. Diz-se que tais genes estão ligados entre si em virtude de estarem localizados no mesmo cromossoma. O número de grupos de genes ligados é o mesmo que o número de cromossomas (quatro em Drosophila), suportando a ideia de que os cromossomas são portadores dos genes.

Figure 3.3
Segregação e ligação dos genes. (A) Segregação de dois genes hipotéticos para forma (A/a = quadrado/redondo) e cor (B/b = vermelho/azul) localizados em cromossomos diferentes. (B) Ligação de dois genes localizados no mesmo cromossoma.
Ligação entre genes não está completa, entretanto; cromossomos trocam material durante a meiose, levando à recombinação entre genes ligados (Figura 3.4). A frequência de recombinação entre dois genes ligados depende da distância entre eles no cromossomo; genes que estão próximos um do outro recombinam com menos frequência do que genes mais distantes. Assim, as frequências com que os diferentes genes se recombinam podem ser usadas para determinar suas posições relativas no cromossomo, permitindo a construção de mapas genéticos (Figura 3.5). Em 1915, quase uma centena de genes havia sido definida e mapeada nos quatro cromossomos de Drosophila, levando à aceitação geral da base cromossômica de hereditariedade.
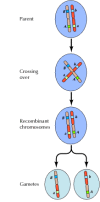
Figure 3.4
Recombinação genética. Durante a meiose, os membros dos pares de cromossomas trocam material. O resultado é a recombinação entre os genes ligados.
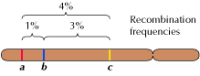
Figura 3.5
Um mapa genético. Três genes são localizados em um cromossomo hipotético baseado em freqüências de recombinação entre eles (1% de recombinação entre a e b; 3% entre b e c; 4% entre a e c). As freqüências de recombinação são aproximadamente proporcionais (mais…)