Genes y cromosomas
Los principios clásicos de la genética fueron deducidos por Gregor Mendel en 1865, a partir de los resultados de experimentos de cultivo con guisantes. Mendel estudió la herencia de una serie de rasgos bien definidos, como el color de las semillas, y pudo deducir reglas generales para su transmisión. En todos los casos, pudo interpretar correctamente los patrones de herencia observados asumiendo que cada rasgo está determinado por un par de factores hereditarios, que ahora se denominan genes. Una copia del gen (llamada alelo) que especifica cada rasgo se hereda de cada progenitor. Por ejemplo, la reproducción de dos cepas de guisantes -una con semillas amarillas y otra con semillas verdes- da los siguientes resultados (Figura 3.1). Las cepas parentales tienen cada una dos copias idénticas del gen que especifica las semillas amarillas (Y) o verdes (y), respectivamente. Las plantas progenitoras son, por tanto, híbridos que han heredado un gen para las semillas amarillas (Y) y otro para las verdes (y). Todas estas plantas progenitoras (la primera generación filial o F1) tienen semillas amarillas, por lo que se dice que el amarillo (Y) es dominante y el verde (y) recesivo. El genotipo (composición genética) de los guisantes de la F1 es, pues, Yy, y su fenotipo (aspecto físico) es amarillo. Si una descendencia F1 se cruza con otra, dando lugar a una progenie F2, los genes de las semillas amarillas y verdes segregan de forma característica, de manera que la proporción entre las plantas F2 con semillas amarillas y las que tienen semillas verdes es de 3:1.
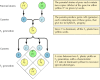
Figura 3.1
Herencia de genes dominantes y recesivos.
Los descubrimientos de Mendel, aparentemente adelantados a su tiempo, fueron ignorados en gran medida hasta 1900, cuando se redescubrieron las leyes de Mendel y se reconoció su importancia. Poco después, se propuso el papel de los cromosomas como portadores de genes. Se descubrió que la mayoría de las células de las plantas superiores y de los animales son diploides, es decir, que contienen dos copias de cada cromosoma. Sin embargo, la formación de las células germinales (el esperma y el óvulo) implica un tipo único de división celular (meiosis) en el que sólo se transmite un miembro de cada par de cromosomas a cada célula de la progenie (figura 3.2). Por consiguiente, el espermatozoide y el óvulo son haploides y sólo contienen una copia de cada cromosoma. La unión de estas dos células haploides en la fecundación crea un nuevo organismo diploide, que ahora contiene un miembro de cada par cromosómico derivado del progenitor masculino y otro del femenino. El comportamiento de los pares de cromosomas es, por tanto, paralelo al de los genes, lo que lleva a la conclusión de que los genes son portados por los cromosomas.
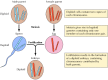
Figura 3.2
Cromosomas en la meiosis y la fecundación. Se ilustran dos pares de cromosomas de un organismo hipotético.
Los fundamentos de la mutación, el ligamiento genético y las relaciones entre los genes y los cromosomas se establecieron en gran medida gracias a los experimentos realizados con la mosca de la fruta, Drosophila melanogaster. La Drosophila puede mantenerse fácilmente en el laboratorio y se reproduce aproximadamente cada dos semanas, lo que supone una ventaja considerable para los experimentos genéticos. De hecho, estas características siguen haciendo de la Drosophila un organismo de elección para los estudios genéticos de los animales, en particular el análisis genético del desarrollo y la diferenciación.
A principios de la década de 1900, se identificaron una serie de alteraciones genéticas (mutaciones) en la Drosophila, que normalmente afectaban a características fácilmente observables como el color de los ojos o la forma de las alas. Los experimentos de cría indicaron que algunos de los genes que gobiernan estos rasgos se heredan de forma independiente, lo que sugiere que estos genes están localizados en diferentes cromosomas que se segregan de forma independiente durante la meiosis (Figura 3.3). Otros genes, sin embargo, se heredan con frecuencia juntos como rasgos emparejados. Se dice que estos genes están vinculados entre sí por estar situados en el mismo cromosoma. El número de grupos de genes ligados es el mismo que el número de cromosomas (cuatro en Drosophila), lo que apoya la idea de que los cromosomas son portadores de los genes.

Figura 3.3
Segregación y ligamiento de genes. (A) Segregación de dos genes hipotéticos para la forma (A/a = cuadrado/redondo) y el color (B/b = rojo/azul) localizados en diferentes cromosomas. (B) Vinculación de dos genes situados en el mismo cromosoma.
El enlace entre genes no es completo, sin embargo; los cromosomas intercambian material durante la meiosis, dando lugar a la recombinación entre los genes enlazados (Figura 3.4). La frecuencia de recombinación entre dos genes enlazados depende de la distancia entre ellos en el cromosoma; los genes que están cerca uno del otro se recombinan con menos frecuencia que los genes más alejados. Así, las frecuencias con las que se recombinan diferentes genes pueden utilizarse para determinar sus posiciones relativas en el cromosoma, lo que permite la construcción de mapas genéticos (figura 3.5). En 1915, se habían definido y cartografiado casi un centenar de genes en los cuatro cromosomas de Drosophila, lo que condujo a la aceptación general de la base cromosómica de la herencia.
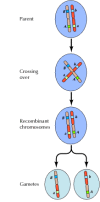
Figura 3.4
Recombinación genética. Durante la meiosis, los miembros de los pares de cromosomas intercambian material. El resultado es la recombinación entre genes ligados.
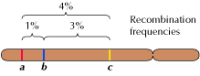
Figura 3.5
Un mapa genético. Se localizan tres genes en un cromosoma hipotético basándose en las frecuencias de recombinación entre ellos (1% de recombinación entre a y b; 3% entre b y c; 4% entre a y c). Las frecuencias de recombinación son aproximadamente proporcionales (más…)