Gener og kromosomer
De klassiske principper for genetik blev udledt af Gregor Mendel i 1865 på baggrund af resultaterne af avlsforsøg med ærter. Mendel studerede arven af en række veldefinerede egenskaber, f.eks. frøfarve, og kunne udlede generelle regler for deres overførsel. I alle tilfælde kunne han fortolke de observerede arvelighedsmønstre korrekt ved at antage, at hver egenskab bestemmes af et par arvelige faktorer, som nu kaldes gener. En genkopi (kaldet en allel), der specificerer hver egenskab, nedarves fra hver forælder. Hvis man f.eks. avler to ærtestammer, hvoraf den ene har gule frø, og den anden grønne frø, får man følgende resultater (figur 3.1). Forældrestammerne har hver to identiske kopier af det gen, der specificerer henholdsvis gule (Y) eller grønne (y) frø. Afkomsplanterne er derfor hybrider, der har arvet et gen for gule frø (Y) og et gen for grønne frø (y). Alle disse afkomsplanter (den første stamme- eller F1-generation) har gule frø, så gult (Y) siges at være dominerende og grønt (y) recessivt. Genotypen (den genetiske sammensætning) af F1-ærterne er således Yy, og deres fænotype (fysisk udseende) er gul. Hvis et F1-afkom avles med et andet, hvorved der opstår F2-afkom, segregeres generne for gule og grønne frø på en karakteristisk måde, således at forholdet mellem F2-planter med gule frø og planter med grønne frø er 3:1.
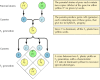
Figur 3.1
Arv af dominerende og recessive gener.
Mendels resultater, der tilsyneladende var forud for deres tid, blev stort set ignoreret indtil 1900, hvor Mendels love blev genopdaget, og deres betydning blev anerkendt. Kort tid efter blev kromosomernes rolle som bærere af generne foreslået. Man indså, at de fleste celler i højere planter og dyr er diploide – de indeholder to kopier af hvert kromosom. Dannelsen af kønscellerne (sædcellerne og ægget) involverer imidlertid en unik form for celledeling (meiose), hvor kun ét medlem af hvert kromosompar overføres til hver afkomscelle (figur 3.2). Derfor er sædcellerne og ægget haploide, dvs. de indeholder kun én kopi af hvert kromosom. Foreningen af disse to haploide celler ved befrugtning skaber en ny diploid organisme, som nu indeholder et medlem af hvert kromosompar, der stammer fra den mandlige og et fra den kvindelige forælder. Kromosomparenes adfærd er således parallel til genernes, hvilket fører til den konklusion, at generne bæres på kromosomer.
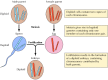
Figur 3.2
Kromosomer ved meiose og befrugtning. To kromosompar fra en hypotetisk organisme er illustreret.
De grundlæggende principper for mutation, genetisk binding og forholdet mellem gener og kromosomer blev i vid udstrækning fastlagt ved forsøg udført med frugtfluen, Drosophila melanogaster. Drosophila kan let vedligeholdes i laboratoriet, og de formerer sig ca. hver anden uge, hvilket er en betydelig fordel for genetiske eksperimenter. Disse egenskaber gør fortsat Drosophila til en foretrukken organisme til genetiske undersøgelser af dyr, især genetisk analyse af udvikling og differentiering.
I begyndelsen af 1900-tallet blev der identificeret en række genetiske ændringer (mutationer) i Drosophila, som normalt påvirker let observerbare egenskaber som f.eks. øjenfarve eller vingeform. Avlsforsøg viste, at nogle af de gener, der styrer disse egenskaber, nedarves uafhængigt af hinanden, hvilket tyder på, at disse gener er placeret på forskellige kromosomer, der segregeres uafhængigt af hinanden under meiosen (figur 3.3). Andre gener nedarves imidlertid ofte sammen som parvise egenskaber. Sådanne gener siges at være knyttet til hinanden i kraft af at være placeret på det samme kromosom. Antallet af grupper af sammenkædede gener er det samme som antallet af kromosomer (fire hos Drosophila), hvilket understøtter tanken om, at kromosomerne er bærere af generne.

Figur 3.3
Gensegregation og sammenkædning. (A) Segregation af to hypotetiske gener for form (A/a = firkantet/rund) og farve (B/b = rød/blå) placeret på forskellige kromosomer. (B) Sammenkædning af to gener, der er placeret på samme kromosom.
Linkage mellem generne er dog ikke fuldstændig; kromosomer udveksler materiale under meiosen, hvilket fører til rekombination mellem de linkede gener (figur 3.4). Hyppigheden af rekombination mellem to forbundne gener afhænger af afstanden mellem dem på kromosomet; gener, der er tæt på hinanden, rekombinerer mindre hyppigt end gener, der er længere fra hinanden. De hyppigheder, hvormed forskellige gener rekombinerer, kan således bruges til at bestemme deres relative positioner på kromosomet, hvilket gør det muligt at konstruere genetiske kort (figur 3.5). I 1915 var næsten hundrede gener blevet defineret og kortlagt på Drosophila’s fire kromosomer, hvilket førte til en generel accept af det kromosomale grundlag for arvelighed.
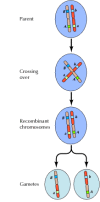
Figur 3.4
Genisk rekombination. Under meiosen udveksler medlemmer af kromosompar materiale. Resultatet er rekombination mellem forbundne gener.
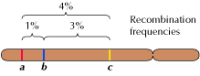
Figur 3.5
Et genetisk kort. Tre gener er lokaliseret på et hypotetisk kromosom baseret på frekvenser af rekombination mellem dem (1 % rekombination mellem a og b; 3 % mellem b og c; 3 % mellem b og c; 4 % mellem a og c). Rekombinationshyppighederne er omtrent proportionale (mere…)