Gene und Chromosomen
Die klassischen Prinzipien der Genetik wurden 1865 von Gregor Mendel auf der Grundlage der Ergebnisse von Züchtungsexperimenten mit Erbsen abgeleitet. Mendel untersuchte die Vererbung einer Reihe genau definierter Merkmale, wie z.B. der Samenfarbe, und konnte daraus allgemeine Regeln für deren Vererbung ableiten. In allen Fällen konnte er die beobachteten Vererbungsmuster korrekt interpretieren, indem er davon ausging, dass jedes Merkmal durch ein Paar von Erbfaktoren, die heute als Gene bezeichnet werden, bestimmt wird. Von jedem Elternteil wird eine Genkopie (ein so genanntes Allel) vererbt, die jedes Merkmal spezifiziert. Wenn man zum Beispiel zwei Erbsenstämme züchtet – einen mit gelben und einen mit grünen Samen -, erhält man folgende Ergebnisse (Abbildung 3.1). Die Elternstämme haben jeweils zwei identische Kopien des Gens, das gelbe (Y) bzw. grüne (y) Samen spezifiziert. Die Nachkommen sind also Hybride, die ein Gen für gelbe Samen (Y) und eines für grüne Samen (y) geerbt haben. Alle diese Nachkommen (die erste Filialgeneration oder F1) haben gelbe Samen, so dass man sagt, dass gelb (Y) dominant und grün (y) rezessiv ist. Der Genotyp (genetische Zusammensetzung) der F1-Erbsen ist also Yy, und ihr Phänotyp (physisches Aussehen) ist gelb. Wenn ein F1-Nachkomme mit einem anderen gekreuzt wird, wodurch F2-Nachkommen entstehen, segregieren die Gene für gelbe und grüne Samen in einer charakteristischen Weise, so dass das Verhältnis zwischen F2-Pflanzen mit gelben Samen und solchen mit grünen Samen 3:1 ist.
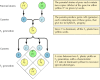
Abbildung 3.1
Vererbung von dominanten und rezessiven Genen.
Mendels Erkenntnisse, die offensichtlich ihrer Zeit voraus waren, wurden bis 1900 weitgehend ignoriert, als die Mendelschen Gesetze wiederentdeckt und ihre Bedeutung erkannt wurde. Kurz darauf wurde die Rolle der Chromosomen als Träger von Genen vorgeschlagen. Man erkannte, dass die meisten Zellen höherer Pflanzen und Tiere diploid sind – sie enthalten zwei Kopien jedes Chromosoms. Bei der Bildung der Keimzellen (Spermien und Eizellen) findet jedoch eine besondere Art der Zellteilung (Meiose) statt, bei der nur ein Mitglied jedes Chromosomenpaars an jede Nachkommenzelle weitergegeben wird (Abbildung 3.2). Folglich sind Spermien und Eizellen haploid, d. h. sie enthalten nur eine Kopie jedes Chromosoms. Durch die Vereinigung dieser beiden haploiden Zellen bei der Befruchtung entsteht ein neuer diploider Organismus, der nun ein Mitglied jedes Chromosomenpaares enthält, das vom männlichen und eines vom weiblichen Elternteil stammt. Das Verhalten von Chromosomenpaaren entspricht also dem von Genen, was zu der Schlussfolgerung führt, dass Gene auf Chromosomen getragen werden.
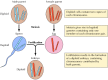
Abbildung 3.2
Chromosomen bei Meiose und Befruchtung. Dargestellt sind zwei Chromosomenpaare eines hypothetischen Organismus.
Die Grundlagen der Mutation, der genetischen Verknüpfung und der Beziehungen zwischen Genen und Chromosomen wurden weitgehend durch Experimente mit der Fruchtfliege Drosophila melanogaster geschaffen. Drosophila kann im Labor leicht gehalten werden und pflanzt sich etwa alle zwei Wochen fort, was für genetische Experimente einen erheblichen Vorteil darstellt. Diese Eigenschaften machen Drosophila auch heute noch zum bevorzugten Organismus für genetische Studien an Tieren, insbesondere für die genetische Analyse von Entwicklung und Differenzierung.
In den frühen 1900er Jahren wurden bei Drosophila eine Reihe von genetischen Veränderungen (Mutationen) festgestellt, die in der Regel leicht zu beobachtende Merkmale wie die Augenfarbe oder die Flügelform betrafen. Zuchtexperimente zeigten, dass einige der Gene, die für diese Merkmale verantwortlich sind, unabhängig voneinander vererbt werden, was darauf hindeutet, dass sich diese Gene auf verschiedenen Chromosomen befinden, die sich während der Meiose unabhängig voneinander segregieren (Abbildung 3.3). Andere Gene werden jedoch häufig zusammen als gepaarte Merkmale vererbt. Solche Gene werden als miteinander verbunden bezeichnet, weil sie sich auf demselben Chromosom befinden. Die Anzahl der Gruppen miteinander verbundener Gene entspricht der Anzahl der Chromosomen (vier bei Drosophila), was die Idee unterstützt, dass die Chromosomen Träger der Gene sind.

Abbildung 3.3
Gen-Segregation und Kopplung. (A) Segregation von zwei hypothetischen Genen für Form (A/a = quadratisch/rund) und Farbe (B/b = rot/blau), die auf verschiedenen Chromosomen liegen. (B) Verknüpfung von zwei Genen, die sich auf demselben Chromosom befinden.
Die Verknüpfung zwischen den Genen ist jedoch nicht vollständig; Chromosomen tauschen während der Meiose Material aus, was zu einer Rekombination zwischen verknüpften Genen führt (Abbildung 3.4). Die Häufigkeit der Rekombination zwischen zwei verknüpften Genen hängt vom Abstand zwischen ihnen auf dem Chromosom ab; Gene, die nahe beieinander liegen, rekombinieren weniger häufig als Gene, die weiter voneinander entfernt sind. Anhand der Häufigkeit, mit der sich verschiedene Gene rekombinieren, lässt sich also ihre relative Position auf dem Chromosom bestimmen, was die Erstellung genetischer Karten ermöglicht (Abbildung 3.5). Bis 1915 waren fast hundert Gene definiert und auf den vier Chromosomen von Drosophila kartiert worden, was zur allgemeinen Anerkennung der chromosomalen Grundlage der Vererbung führte.
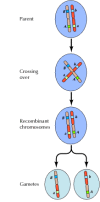
Abbildung 3.4
Genetische Rekombination. Während der Meiose tauschen die Mitglieder von Chromosomenpaaren Material aus. Das Ergebnis ist eine Rekombination zwischen verbundenen Genen.
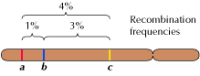
Abbildung 3.5
Eine genetische Karte. Drei Gene sind auf einem hypothetischen Chromosom auf der Grundlage der Rekombinationshäufigkeiten zwischen ihnen lokalisiert (1% Rekombination zwischen a und b; 3% zwischen b und c; 4% zwischen a und c). Die Häufigkeit der Rekombination ist ungefähr proportional (mehr…)