Die DNA-Replikation, auch als semikonservative Replikation bekannt, ist der Prozess, durch den die DNA im Wesentlichen verdoppelt wird. Es ist ein wichtiger Prozess, der in der sich teilenden Zelle stattfindet.
In diesem Artikel werden wir kurz die Struktur der DNA, die genauen Schritte der DNA-Replikation (Initiation, Elongation und Terminierung) und die klinischen Folgen, die auftreten können, wenn dies schief geht, betrachten.
DNA-Struktur
DNA besteht aus Millionen von Nukleotiden. Das sind Moleküle, die aus einem Desoxyribose-Zucker bestehen, an den ein Phosphat und eine Base (oder Nukleobase) gebunden sind. Diese Nukleotide sind in Strängen über Phosphodiesterbindungen miteinander verbunden und bilden ein „Zucker-Phosphat-Grundgerüst“. Die Bindung entsteht zwischen dem dritten Kohlenstoffatom des Desoxyribosezuckers eines Nukleotids (im Folgenden als 3′ bezeichnet) und dem fünften Kohlenstoffatom eines anderen Zuckers des nächsten Nukleotids (als 5′ bezeichnet).
N.B: 3′ wird als ‚drei Primzahlen‘ und 5′ als ‚fünf Primzahlen‘ ausgesprochen.
Es gibt zwei Stränge, die in entgegengesetzter oder antiparalleler Richtung zueinander verlaufen. Diese sind über die gesamte Länge des Strangs durch die Basen an jedem Nukleotid miteinander verbunden. Es gibt 4 verschiedene Basen in der DNA: Cytosin, Guanin, Adenin und Thymin. In normalen DNA-Strängen bindet sich Cytosin an Guanin und Adenin an Thymin. Die beiden Stränge bilden zusammen eine Doppelhelix.
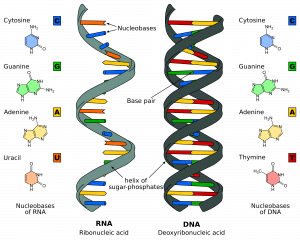 Abb. 1.0 – Die Struktur von RNS und DNS
Abb. 1.0 – Die Struktur von RNS und DNSStadien der DNS-Replikation
Die DNS-Replikation kann in drei Stadien unterteilt werden: Initiation, Elongation, Termination
Initiation
Die DNS-Synthese wird an bestimmten Punkten innerhalb des DNS-Strangs eingeleitet, die als „Origins“ bekannt sind, bei denen es sich um spezifische kodierende Regionen handelt. Diese Ursprünge werden von Initiatorproteinen angesteuert, die dann weitere Proteine rekrutieren, die den Replikationsprozess unterstützen und einen Replikationskomplex um den DNA-Ursprung herum bilden. Es gibt mehrere Ursprungsstellen, und wenn die Replikation der DNA beginnt, werden diese Stellen als Replikationsgabeln bezeichnet.
Innerhalb des Replikationskomplexes befindet sich das Enzym DNA-Helicase, das die Doppelhelix abwickelt und jeden der beiden Stränge freilegt, so dass sie als Vorlage für die Replikation verwendet werden können. Dazu hydrolysiert es das ATP, das zur Bildung der Bindungen zwischen den Nukleobasen verwendet wird, und bricht so die Bindung, die die beiden Stränge zusammenhält.
DNA-Primase ist ein weiteres Enzym, das für die DNA-Replikation wichtig ist. Es synthetisiert einen kleinen RNA-Primer, der als „Kickstarter“ für die DNA-Polymerase dient. Die DNA-Polymerase ist das Enzym, das letztlich für die Bildung und Ausdehnung der neuen DNA-Stränge verantwortlich ist.
Verlängerung
Sobald die DNA-Polymerase sich an die ursprünglichen, noch nicht entpackten zwei DNA-Stränge (d.h. die Vorlagenstränge) angehängt hat, kann sie mit der Synthese der neuen DNA beginnen, die den Vorlagen entspricht. Es ist wichtig zu wissen, dass die DNA-Polymerase den Primer nur verlängern kann, indem sie freie Nukleotide an das 3′-Ende anhängt.
Eine der Vorlagen wird in 3′-5′-Richtung gelesen, was bedeutet, dass der neue Strang in 5′-3′-Richtung gebildet wird. Dieser neu gebildete Strang wird als Führungsstrang bezeichnet. Entlang dieses Strangs muss die DNA-Primase nur einmal, nämlich am Anfang, einen RNA-Primer synthetisieren, um die DNA-Polymerase zu starten. Dies liegt daran, dass die DNA-Polymerase in der Lage ist, den neuen DNA-Strang zu verlängern, indem sie die Vorlage von 3′ nach 5′ liest und wie oben erwähnt in einer Richtung von 5′ nach 3′ synthetisiert.
Der andere Vorlagenstrang (der nachlaufende Strang) ist jedoch antiparallel und wird daher in einer Richtung von 5′ nach 3′ gelesen. Eine kontinuierliche DNA-Synthese wie beim Vorlaufstrang müsste in der Richtung 3′ zu 5′ erfolgen, was unmöglich ist, da wir am 5′-Ende keine Basen hinzufügen können. Stattdessen werden beim Abwickeln der Helix RNA-Primer an die neu freiliegenden Basen des nacheilenden Strangs angehängt, und die DNA-Synthese erfolgt in Fragmenten, aber immer noch in der Richtung 5′ zu 3′ wie zuvor. Diese Fragmente werden als Okazaki-Fragmente bezeichnet.
Termination
Der Prozess der Expansion der neuen DNA-Stränge wird so lange fortgesetzt, bis entweder keine DNA-Vorlage mehr für die Replikation vorhanden ist (d. h. am Ende des Chromosoms) oder zwei Replikationsgabeln aufeinandertreffen und anschließend terminieren. Das Zusammentreffen zweier Replikationsgabeln ist nicht reguliert und geschieht zufällig im Verlauf des Chromosoms.
Nach Abschluss der DNA-Synthese ist es wichtig, dass die neu synthetisierten Stränge gebunden und stabilisiert werden. Dazu werden zwei Enzyme benötigt: RNAase H entfernt den RNA-Primer, der sich am Anfang jedes Okazaki-Fragments befindet, und DNA-Ligase fügt die Fragmente zu einem vollständigen Strang zusammen.
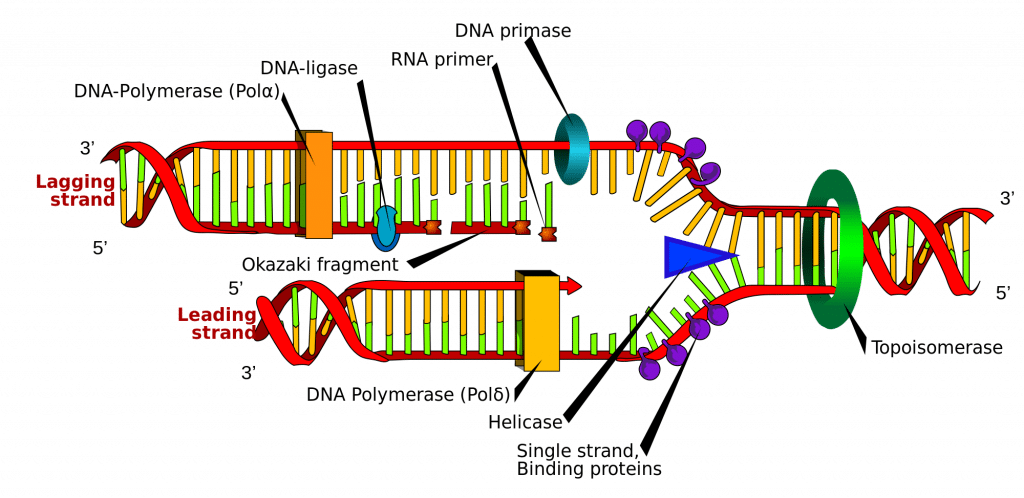 Abb. 2.0 – Schematische Darstellung der DNA-Replikation
Abb. 2.0 – Schematische Darstellung der DNA-Replikation
Klinische Relevanz – Sichelzellenanämie
Die Sichelzellenanämie ist eine autosomal rezessiv vererbte Erkrankung, die durch eine einzige Basensubstitution verursacht wird, bei der nur eine Base gegen eine andere ausgetauscht wird. In einigen Fällen kann dies zu einer „stummen Mutation“ führen, bei der das gesamte Gen nicht betroffen ist; bei Krankheiten wie der Sichelzellenanämie führt es jedoch dazu, dass der Strang für ein anderes Protein kodiert.
In diesem Fall wird in einem der Gene, die für Hämoglobin kodieren, eine Adeninbase gegen eine Thyminbase ausgetauscht; dies führt dazu, dass Glutaminsäure durch Valin ersetzt wird. Bei der Transkription in eine Polypeptidkette ändern sich deren Eigenschaften radikal, da Glutaminsäure hydrophil ist, während Valin hydrophob ist. Diese hydrophobe Region führt dazu, dass Hämoglobin eine anormale Struktur aufweist, die zu Verstopfungen von Kapillaren führen kann, was eine Ischämie und möglicherweise eine Nekrose von Geweben und Organen zur Folge hat – dies wird als vasookklusive Krise bezeichnet.
Diese Krisen werden in der Regel mit einer Reihe von Schmerzmitteln, einschließlich Opioiden und NSAIDs, je nach Schweregrad behandelt. In Notfällen können Transfusionen von roten Blutkörperchen erforderlich sein, zum Beispiel wenn die Blockade in der Lunge auftritt.
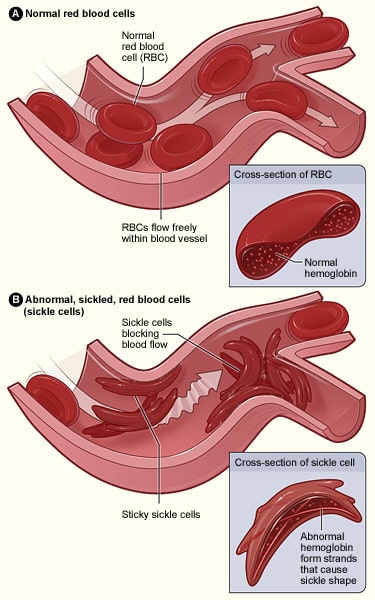 Abb. 3.0 – Der Unterschied in der Struktur zwischen normalen roten Blutkörperchen und solchen, die von der Sichelzellkrankheit betroffen sind.
Abb. 3.0 – Der Unterschied in der Struktur zwischen normalen roten Blutkörperchen und solchen, die von der Sichelzellkrankheit betroffen sind.