DNA:n replikaatio, joka tunnetaan myös nimellä puolikonservatiivinen replikaatio, on prosessi, jonka avulla DNA:ta olennaisesti kaksinkertaistetaan. Se on tärkeä prosessi, joka tapahtuu jakautuvassa solussa.
Tässä artikkelissa tarkastelemme lyhyesti DNA:n rakennetta, DNA:n replikaatioon liittyviä täsmällisiä vaiheita (initiaatio, pidennys ja lopetus) sekä kliinisiä seurauksia, joita voi syntyä, kun replikaatio menee pieleen.
DNA:n rakenne
DNA:n rakenne
DNA:n rakenne koostuu miljoonista nukleotideista. Nämä ovat molekyylejä, jotka koostuvat deoksiriboosisokerista, johon on liitetty fosfaatti ja emäs (tai nukleobaasi). Nämä nukleotidit ovat kiinnittyneet toisiinsa säikeiksi fosfodiesterisidoksilla muodostaen ”sokeri-fosfaattirungon”. Muodostuva sidos muodostuu yhden nukleotidin deoksiriboosisokerin kolmannen hiiliatomin (jäljempänä 3′) ja seuraavan nukleotidin toisen sokerin viidennen hiiliatomin (5′) välille.
N.B.: 3′ lausutaan ’kolme priimaa’ ja 5′ lausutaan ’viisi priimaa’.
Kaksi säiettä kulkevat vastakkaisiin tai antiparalleelisiin suuntiin toisiinsa nähden. Ne ovat kiinnittyneet toisiinsa koko säikeen pituudelta kunkin nukleotidin emästen välityksellä. DNA:han liittyy neljä eri emästä; sytosiini, guaniini, adeniini ja tymiini. Normaaleissa DNA-juosteissa sytosiini sitoutuu guaniiniin ja adeniini tymiiniin. Nämä kaksi säiettä muodostavat yhdessä kaksoiskierteen.
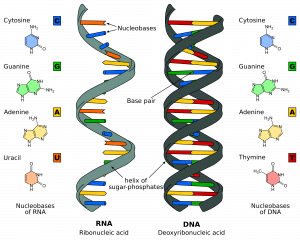 Kuva 1.0 – RNA:n ja DNA:n rakenne
Kuva 1.0 – RNA:n ja DNA:n rakenneDNA:n replikaation vaiheet
DNA:n replikaation voidaan ajatella koostuvan kolmesta vaiheesta; aloituksesta, pidentymisestä ja lopetuksesta
Aloituksesta
DNA:n synteesi aloitetaan DNA:n tiettyihin DNA:n sisällä oleviin kohtiin, niin sanottuihin alkupisteisiin (origineihin). Nämä kohdat ovat tietyt koodaavasti koodaavat alueet. Aloittajaproteiinit kohdistuvat näihin alkuperiin, jotka rekrytoivat lisää proteiineja, jotka auttavat replikaatioprosessia ja muodostavat replikaatiokompleksin DNA-alkuperän ympärille. Alkupisteitä on useita, ja kun DNA:n replikaatio alkaa, näitä paikkoja kutsutaan replikaatiohaarukoiksi.
Replikaatiokompleksissa on entsyymi DNA-helikaasi, joka purkaa kaksoiskierteen ja paljastaa molemmat säikeet, jotta niitä voidaan käyttää mallina replikaatiossa. Se tekee tämän hydrolysoimalla ATP:tä, jota on käytetty nukleoaasien välisten sidosten muodostamiseen, ja katkaisee siten sidoksen, joka pitää kaksi säiettä yhdessä.
DNA-primaasi on toinen entsyymi, joka on tärkeä DNA:n replikaatiossa. Se syntetisoi pienen RNA-alukkeen, joka toimii DNA-polymeraasin ”käynnistäjänä”. DNA-polymeraasi on entsyymi, joka on viime kädessä vastuussa uusien DNA-säikeiden luomisesta ja laajentamisesta.
Pidennys
Kun DNA-polymeraasi on kiinnittynyt alkuperäisiin, purettuihin kahteen DNA-säikeeseen (eli templaattisäikeisiin), se pystyy aloittamaan uuden DNA:n syntetisoinnin vastaamaan malleja. On tärkeää huomata, että DNA-polymeraasi pystyy pidentämään aluketta vain lisäämällä vapaita nukleotideja 3′-päähän.
Jompikumpi templaateista luetaan 3′-5′-suunnassa, mikä tarkoittaa, että uusi säie muodostetaan 5′-3′-suunnassa. Tätä uutta muodostunutta säiettä kutsutaan johtavaksi säikeeksi. Tätä säiettä pitkin DNA-primaasin tarvitsee syntetisoida RNA-alkuaine vain kerran, alussa, DNA-polymeraasin käynnistämiseksi. Tämä johtuu siitä, että DNA-polymeraasi pystyy pidentämään uutta DNA-juostetta lukemalla templaattia 3′:sta 5′:een ja syntetisoimalla sen 5′:sta 3′:een suuntaan, kuten edellä todettiin.
Mutta toinen templaattisäie (jäljessä oleva säie) on antiparalleelinen, joten se luetaan 5′:sta 3′:een suuntaan. Jatkuvan DNA-synteesin, kuten johtavan säikeen, täytyisi tapahtua 3′-5′-suunnassa, mikä on mahdotonta, koska emme voi lisätä emäksiä 5′-päähän. Sen sijaan spiraalin purkautuessa RNA-alukkeet lisätään vasta paljastuneisiin emäksiin viivejuosteessa, ja DNA-synteesi tapahtuu pätkittäin, mutta edelleen 5′-3′-suunnassa kuten aiemmin. Näitä fragmentteja kutsutaan Okazaki-fragmenteiksi.
Terminaatio
Uusien DNA-säikeiden laajenemisprosessi jatkuu, kunnes joko DNA-mallia ei ole enää jäljellä replikoitavaksi (eli kromosomin lopussa) tai kaksi replikaatiohaarukkaa kohtaavat toisensa ja sen jälkeen ne päättyvät. Kahden replikaatiohaarukan kohtaamista ei säädellä, vaan se tapahtuu sattumanvaraisesti kromosomin kulussa.
Kun DNA-synteesi on päättynyt, on tärkeää, että vastasyntetisoidut säikeet sidotaan ja stabiloidaan. Jäljellä olevan säikeen osalta tähän tarvitaan kaksi entsyymiä; RNAaasi H poistaa jokaisen Okazaki-fragmentin alussa olevan RNA-alukkeen, ja DNA-ligaasi yhdistää fragmentit toisiinsa yhdeksi täydelliseksi säikeeksi.
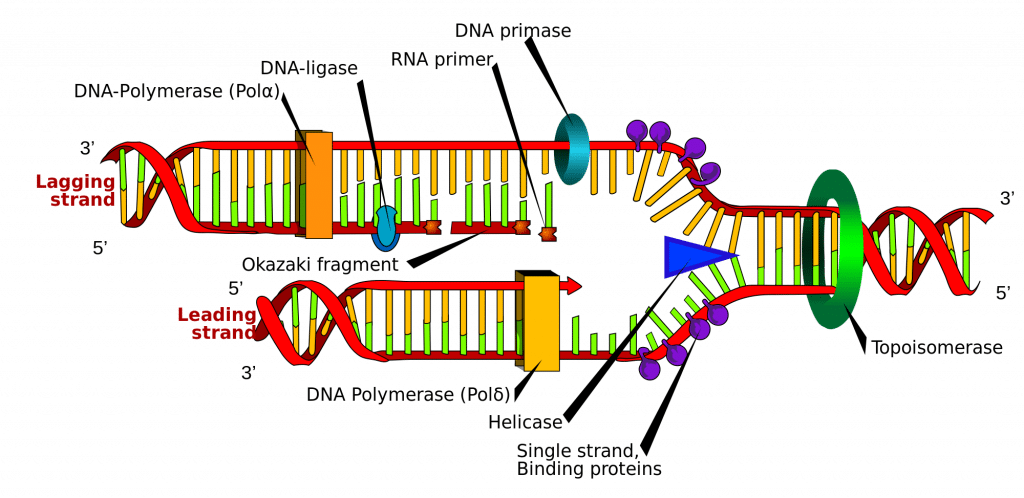 Kuva 2. DNA:n ja DNA:n yhdistäminen.0 – Kaavamainen esitys DNA:n replikaatiosta
Kuva 2. DNA:n ja DNA:n yhdistäminen.0 – Kaavamainen esitys DNA:n replikaatiosta
Kliininen merkitys – Sirppisoluanemia
Sirppisoluanemia on autosomaalinen resessiivinen sairaus, joka johtuu yhden emäksen substituutiosta, jossa vain yksi emäs vaihtuu toiseen. Joissakin tapauksissa tämä voi johtaa ”hiljaiseen mutaatioon”, jossa koko geeni ei vaikuta, mutta sirppisoluanemian kaltaisissa sairauksissa se johtaa siihen, että säie koodaa erilaista proteiinia.
Tässä tapauksessa adeniini-emäs on vaihdettu tymiini-emäkselle yhdessä hemoglobiinia koodaavassa geenissä; tämä johtaa siihen, että glutamiinihappo korvataan valiinilla. Kun tämä kirjoitetaan polypeptidiketjuksi, sen ominaisuudet muuttuvat radikaalisti, koska glutamiinihappo on hydrofiilinen, kun taas valiini on hydrofobinen. Tämä hydrofobinen alue johtaa siihen, että hemoglobiinilla on epänormaali rakenne, joka voi aiheuttaa kapillaarien tukkeutumista, mikä johtaa kudosten ja elinten iskemiaan ja mahdollisesti nekroosiin – tätä kutsutaan vaso-okklusiiviseksi kriisiksi.
Näitä kriisejä hoidetaan tyypillisesti erilaisilla kipulääkkeillä, mukaan lukien opioideilla ja tulehduskipulääkkeillä niiden vakavuudesta riippuen. Punasolusiirtoja saatetaan tarvita hätätilanteissa, esimerkiksi jos tukos esiintyy keuhkoissa.
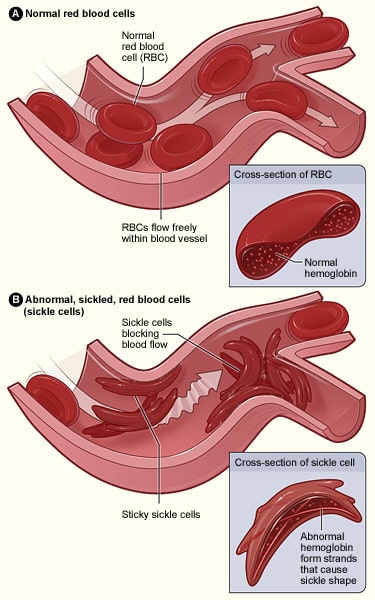 Kuva 3.0 – Normaalien punasolujen ja sirppisolusairauden aiheuttamien punasolujen rakenteen ero.
Kuva 3.0 – Normaalien punasolujen ja sirppisolusairauden aiheuttamien punasolujen rakenteen ero.