Geenit ja kromosomit
Klassiset perinnöllisyysperiaatteet päätteli Gregor Mendel vuonna 1865 herneillä tehtyjen jalostuskokeiden tulosten perusteella. Mendel tutki useiden tarkoin määriteltyjen ominaisuuksien, kuten siementen värin, periytymistä ja pystyi päättelemään yleiset säännöt niiden periytymisestä. Kaikissa tapauksissa hän pystyi tulkitsemaan havaitut periytymismallit oikein olettamalla, että kukin ominaisuus määräytyy parin perintötekijän perusteella, joita nyt kutsutaan geeneiksi. Kummaltakin vanhemmalta periytyy yksi geenikopio (jota kutsutaan alleeliksi), joka määrittää kunkin ominaisuuden. Jos esimerkiksi jalostetaan kahta herneenlajiketta, joista toisella on keltaiset ja toisella vihreät siemenet, saadaan seuraavat tulokset (kuva 3.1). Vanhempaiskannoilla on kummallakin kaksi identtistä kopiota geenistä, joka määrittelee keltaiset (Y) tai vihreät (y) siemenet. Jälkeläiskasvit ovat siis hybridejä, jotka ovat perineet yhden geenin keltaisia siemeniä (Y) ja toisen vihreitä siemeniä (y) varten. Kaikilla näillä jälkeläiskasveilla (ensimmäinen jälkeläissukupolvi eli F1-sukupolvi) on keltaisia siemeniä, joten keltaisen (Y) sanotaan olevan dominoiva ja vihreän (y) resessiivinen. F1-herneiden genotyyppi (geneettinen koostumus) on siis Yy, ja niiden fenotyyppi (fyysinen ulkonäkö) on keltainen. Jos yksi F1- jälkeläinen jalostetaan toisen kanssa, jolloin syntyy F2- jälkeläisiä, keltaisten ja vihreiden siementen geenit segregoituvat tyypillisellä tavalla siten, että keltaisia siemeniä omaavien ja vihreitä siemeniä omaavien F2-kasvien suhde on 3:1.
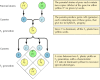
Kuvio 3.1
Herneen dominoivien ja resessiivisten geenien periytyminen.
Mendelin havainnot, jotka olivat ilmeisesti aikaansa edellä, jäivät suurelta osin huomiotta vuoteen 1900 asti, jolloin Mendelin lait löydettiin uudelleen ja niiden merkitys tunnustettiin. Pian tämän jälkeen ehdotettiin kromosomien roolia geenien kantajina. Tajuttiin, että useimmat korkeampien kasvien ja eläinten solut ovat diploideja, eli niissä on kaksi kappaletta kutakin kromosomia. Sukusolujen (siittiöiden ja munasolujen) muodostumiseen liittyy kuitenkin ainutlaatuinen solunjakautuminen (meioosi), jossa kustakin kromosomiparista vain yksi jäsen siirtyy kuhunkin jälkeläissoluun (kuva 3.2). Näin ollen siittiö ja munasolu ovat haploideja, eli niissä on vain yksi kopio kustakin kromosomista. Näiden kahden haploidisen solun yhdistyminen hedelmöityksessä synnyttää uuden diploidisen organismin, joka sisältää nyt yhden jäsenen kustakin kromosomiparista, joka on peräisin urospuoliselta vanhemmalta ja yhden naaraspuoliselta vanhemmalta. Kromosomiparien käyttäytyminen on näin ollen rinnakkaista geenien käyttäytymisen kanssa, mikä johtaa johtopäätökseen, että geenit kulkevat kromosomeissa.
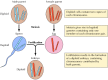
Kuva 3.2
Kromosomit meioosissa ja hedelmöityksessä. Kuvassa on kaksi hypoteettisen organismin kromosomiparia.
Mutaation, geneettisen kytkennän sekä geenien ja kromosomien välisten suhteiden perusteet luotiin suurelta osin hedelmäkärpäsellä, Drosophila melanogasterilla, tehdyissä kokeissa. Drosofilaa on helppo ylläpitää laboratoriossa, ja se lisääntyy noin kahden viikon välein, mikä on huomattava etu geneettisten kokeiden kannalta. Nämä ominaisuudet tekevätkin Drosofilasta edelleen ensisijaisen organismin eläinten geneettisissä tutkimuksissa, erityisesti kehityksen ja erilaistumisen geneettisessä analysoinnissa.
1900-luvun alussa Drosofilasta tunnistettiin useita geneettisiä muutoksia (mutaatioita), jotka yleensä vaikuttivat helposti havaittaviin ominaisuuksiin, kuten silmien väriin tai siipien muotoon. Jalostuskokeet osoittivat, että osa näitä ominaisuuksia säätelevistä geeneistä periytyy toisistaan riippumatta, mikä viittaa siihen, että nämä geenit sijaitsevat eri kromosomeilla, jotka erottuvat itsenäisesti meioosin aikana (kuva 3.3). Toiset geenit kuitenkin periytyvät usein yhdessä parittaisina ominaisuuksina. Tällaisten geenien sanotaan olevan sidoksissa toisiinsa sen vuoksi, että ne sijaitsevat samalla kromosomilla. Linkittyneiden geenien ryhmien lukumäärä on sama kuin kromosomien lukumäärä (neljä Drosophilalla), mikä tukee ajatusta, että kromosomit ovat geenien kantajia.

Kuvio 3.3
Geenien erottuminen ja linkittyminen. (A) Kahden eri kromosomeilla sijaitsevan hypoteettisen muodon (A/a = neliö/pyöreä) ja värin (B/b = punainen/sininen) geenin segregaatio. (B) Samassa kromosomissa sijaitsevien kahden geenin linkittyminen.
Geenien välinen linkittyminen ei kuitenkaan ole täydellistä; kromosomit vaihtavat materiaalia meioosin aikana, mikä johtaa linkittyneiden geenien väliseen rekombinaatioon (kuva 3.4). Kahden linkitetyn geenin välisen rekombinaation taajuus riippuu niiden välisestä etäisyydestä kromosomissa; lähellä toisiaan olevat geenit rekombinoivat harvemmin kuin kauempana toisistaan olevat geenit. Näin ollen eri geenien rekombinaatiotaajuuksia voidaan käyttää niiden suhteellisen sijainnin määrittämiseen kromosomissa, mikä mahdollistaa geneettisten karttojen rakentamisen (kuva 3.5). Vuoteen 1915 mennessä oli määritelty ja kartoitettu lähes sata geeniä Drosophilan neljään kromosomiin, mikä johti siihen, että perinnöllisyyden kromosomaalinen perusta hyväksyttiin yleisesti.
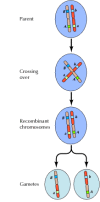
Kuvio 3.4
Geneettinen rekombinaatio. Meioosin aikana kromosomiparien jäsenet vaihtavat materiaalia. Tuloksena on yhdistettyjen geenien välinen rekombinaatio.
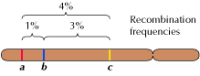
Kuva 3.5
Geneettinen kartta. Kolme geeniä on lokalisoitu hypoteettiselle kromosomille niiden välisen rekombinaation frekvenssien perusteella (1 % rekombinaatio a:n ja b:n välillä; 3 % b:n ja c:n välillä; 4 % a:n ja c:n välillä). Rekombinaatiofrekvenssit ovat suunnilleen verrannollisia (lisää…)