遺伝子と染色体
遺伝学の古典的原理は、1865年にグレゴール・メンデルによって、エンドウの育種実験の結果に基づいて推論されたものです。 メンデルは、種子の色など、明確に定義された形質の遺伝を研究し、その伝達に関する一般法則を導き出すことができた。 どのような場合でも、メンデルは、各形質が現在遺伝子と呼ばれている一対の遺伝因子によって決定されると仮定することによって、観察された遺伝のパターンを正しく解釈することができた。 それぞれの形質を特定する1つの遺伝子コピー(対立遺伝子と呼ばれる)が、それぞれの親から受け継がれるのである。 例えば、黄色い種子を持つエンドウと緑色の種子を持つエンドウの2系統を交配すると、次のような結果になる(図3.1)。 親株はそれぞれ、黄色(Y)または緑色(Y)の種子を指定する遺伝子の同一コピーを2つずつ持っている。 したがって、子孫の植物は、黄色の種子(Y)と緑色の種子(y)の遺伝子を1つずつ受け継いだ雑種である。 この子孫植物(第一孝行世代、F1世代)はすべて黄色の種子を持つので、黄色(Y)は優性、緑色(Y)は劣性と言われる。 したがって、F1エンドウの遺伝子型(遺伝子の構成)はYyであり、その表現型(身体的外観)は黄色である。 F1の子孫と別の子孫を交配してF2の子孫を作ると、黄色と緑の種子の遺伝子は、黄色の種子を持つF2植物と緑の種子を持つものの比率が3:1になるように特徴的に分離する。
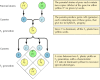
図 3.1
優勢と劣勢の遺伝子を継承する。
メンデルの発見は、明らかに時代を先取りしていたが、1900年にメンデルの法則が再発見されその重要性が認識されるまで、ほとんど無視された。 その後まもなく、遺伝子の運び手としての染色体の役割が提唱された。 高等植物や動物のほとんどの細胞は2倍体であり、各染色体のコピーを2つ持っていることが明らかになった。 しかし、生殖細胞(精子と卵子)の形成には独特の細胞分裂(減数分裂)があり、各染色体対のうち1つだけが子孫の細胞に受け継がれます(図3.2)。 その結果、精子と卵は各染色体のコピーを1つずつしか含まないハプロイドとなる。 受精の際、この2つのハプロイド細胞が結合すると、新しい2倍体の生物が誕生する。このとき、各染色体対の1つは雄から、1つは雌の親から受け継いだものである。
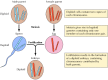
図3.2
減数分裂と受精の際の染色体。 仮想的な生物の2対の染色体が描かれています。
突然変異、遺伝子の連鎖、遺伝子と染色体の関係などの基礎は、ショウジョウバエを使った実験によってほぼ確立されました。 ショウジョウバエは実験室で簡単に飼育でき、約2週間ごとに繁殖するので、遺伝子実験にはかなり有利である。 実際、これらの特徴から、ショウジョウバエは動物の遺伝学的研究、特に発生と分化の遺伝学的分析に適した生物であり続けている。
1900 年代初頭、ショウジョウバエでは多くの遺伝子変化(突然変異)が確認されたが、通常は目の色や翼の形など容易に観察できる特徴に影響を与えるものだった。 繁殖実験の結果、これらの形質を支配する遺伝子のいくつかは互いに独立して遺伝することが示された。これは、これらの遺伝子が異なる染色体上にあり、減数分裂の際に独立して分離することを示唆している(図3.3)。 しかし、他の遺伝子は、対になった特性として一緒に遺伝することが多い。 このような遺伝子は、同じ染色体上に存在することにより、互いにリンクしていると言われています。 このような遺伝子の集団は、染色体の数と同じ数(ショウジョウバエでは4本)であり、染色体が遺伝子のキャリアーであるという考え方が支持されます。 (A)異なる染色体上にある形(A/a=四角/丸)と色(B/b=赤/青)の2つの仮想遺伝子の分離。 (B)同じ染色体上にある2つの遺伝子の連鎖。
ただし、遺伝子間の結合は完全ではなく、減数分裂の際に染色体が交換され、結合した遺伝子間で組換えが起こります(図3.4)。 2つの遺伝子間の組換えは染色体上の距離によって異なり、近い遺伝子は遠い遺伝子より組換え頻度が低い。 このように、異なる遺伝子が組み換わる頻度から、染色体上の相対的な位置を決定することができ、遺伝地図の作成が可能になる(図3.5)。 1915年までには、ショウジョウバエの4本の染色体上に100個近い遺伝子が定義され、マッピングされ、遺伝は染色体に基づいているということが一般に受け入れられるようになった。 減数分裂の際に、染色体対のメンバーが物質を交換します。 その結果、連結した遺伝子間で組換えが起こる。
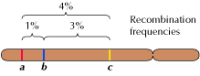
図3.5
遺伝地図。 3つの遺伝子が仮想の染色体上に、それらの間の組換え頻度(aとbの間で1%、bとcの間で3%、aとcの間で4%)をもとに局在している。 組換え頻度はほぼ比例する(以上)
。