Genen en Chromosomen
De klassieke principes van de genetica werden in 1865 door Gregor Mendel afgeleid op basis van de resultaten van kweekexperimenten met erwten. Mendel bestudeerde de vererving van een aantal welomschreven eigenschappen, zoals de kleur van zaden, en kon daaruit algemene regels afleiden voor de overdracht daarvan. In alle gevallen kon hij de waargenomen patronen van overerving correct interpreteren door aan te nemen dat elke eigenschap wordt bepaald door een paar erfelijke factoren, die nu genen worden genoemd. Van elke ouder wordt één genkopie (allel genoemd) geërfd die elke eigenschap specificeert. Bijvoorbeeld, het kweken van twee erwtenrassen – het ene met gele zaden, het andere met groene zaden – levert de volgende resultaten op (figuur 3.1). De ouderstammen hebben elk twee identieke kopieën van het gen dat respectievelijk geel (Y) of groen (y) zaad specificeert. De nateeltplanten zijn dus hybriden, die één gen voor gele zaden (Y) en één voor groene zaden (y) hebben geërfd. Al deze nateeltplanten (de eerste nateelt- of F1-generatie) hebben gele zaden, zodat wordt gezegd dat geel (Y) dominant is en groen (y) recessief. Het genotype (genetische samenstelling) van de F1 erwten is dus Yy, en hun fenotype (fysieke verschijning) is geel. Als een F1-nakomeling met een andere wordt gekruist, waaruit F2-nakomelingen ontstaan, segregeren de genen voor gele en groene zaden op een karakteristieke manier, zodat de verhouding tussen F2-planten met gele zaden en die met groene zaden 3:1 is.
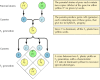
Figuur 3.1
Vererving van dominante en recessieve genen.
Mendel’s bevindingen, die kennelijk hun tijd ver vooruit waren, werden grotendeels genegeerd tot 1900, toen Mendel’s wetten werden herontdekt en hun belang werd erkend. Kort daarna werd de rol van chromosomen als dragers van genen voorgesteld. Men realiseerde zich dat de meeste cellen van hogere planten en dieren diploïd zijn – ze bevatten twee exemplaren van elk chromosoom. Voor de vorming van de kiemcellen (het sperma en de eicel) is echter een unieke vorm van celdeling (meiose) nodig, waarbij slechts één lid van elk chromosomenpaar aan elke nageslachtscel wordt doorgegeven (figuur 3.2). Het sperma en de eicel zijn dus haploïd en bevatten slechts één exemplaar van elk chromosoom. Door de vereniging van deze twee haploïde cellen bij de bevruchting ontstaat een nieuw diploïd organisme, dat nu van elk chromosomenpaar één lid bevat dat afkomstig is van de mannelijke ouder en één van de vrouwelijke ouder. Het gedrag van chromosomenparen komt dus overeen met dat van genen, wat tot de conclusie leidt dat genen op chromosomen worden gedragen.
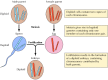
Figuur 3.2
Chromosomen bij meiose en bevruchting. Twee chromosomenparen van een hypothetisch organisme zijn afgebeeld.
De grondbeginselen van mutatie, genetische koppeling en de relaties tussen genen en chromosomen zijn grotendeels vastgesteld door experimenten die zijn uitgevoerd met de fruitvlieg, Drosophila melanogaster. Drosophila kan gemakkelijk in het laboratorium worden gehouden en plant zich ongeveer om de twee weken voort, wat een aanzienlijk voordeel is voor genetische experimenten. Deze eigenschappen maken Drosophila nog steeds tot het favoriete organisme voor genetische studies van dieren, in het bijzonder voor de genetische analyse van ontwikkeling en differentiatie.
In het begin van de 20e eeuw werden bij Drosophila een aantal genetische veranderingen (mutaties) geïdentificeerd, die meestal gemakkelijk waarneembare kenmerken aantasten, zoals de kleur van de ogen of de vorm van de vleugels. Fokexperimenten wezen uit dat sommige van de genen die deze eigenschappen bepalen onafhankelijk van elkaar vererven, wat suggereert dat deze genen zich op verschillende chromosomen bevinden die tijdens de meiose onafhankelijk segregeren (Figuur 3.3). Andere genen daarentegen vererven vaak samen als gepaarde kenmerken. Van dergelijke genen wordt gezegd dat ze aan elkaar gekoppeld zijn omdat ze zich op hetzelfde chromosoom bevinden. Het aantal groepen van gekoppelde genen is even groot als het aantal chromosomen (vier bij Drosophila), wat het idee ondersteunt dat chromosomen dragers van de genen zijn.

Figuur 3.3
Gene segregatie en linkage. (A) Segregatie van twee hypothetische genen voor vorm (A/a = vierkant/rond) en kleur (B/b = rood/blauw) gelegen op verschillende chromosomen. (B) Koppeling van twee genen op hetzelfde chromosoom.
De koppeling tussen genen is echter niet volledig; chromosomen wisselen tijdens de meiose materiaal uit, wat leidt tot recombinatie tussen gekoppelde genen (figuur 3.4). De frequentie van recombinatie tussen twee gekoppelde genen hangt af van de afstand tussen beide op het chromosoom; genen die dicht bij elkaar liggen recombineren minder vaak dan genen die verder van elkaar liggen. De frequentie waarmee verschillende genen recombineren kan dus worden gebruikt om hun relatieve positie op het chromosoom te bepalen, waardoor genetische kaarten kunnen worden gemaakt (figuur 3.5). Tegen 1915 waren bijna honderd genen gedefinieerd en in kaart gebracht op de vier chromosomen van Drosophila, wat leidde tot algemene aanvaarding van de chromosomale basis van erfelijkheid.
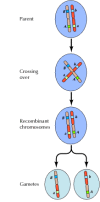
Figuur 3.4
Genetische recombinatie. Tijdens de meiose wisselen leden van chromosomenparen materiaal uit. Het resultaat is recombinatie tussen gekoppelde genen.
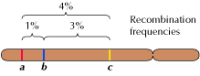
Figuur 3.5
Een genetische kaart. Drie genen zijn gelokaliseerd op een hypothetisch chromosoom op basis van frequenties van recombinatie tussen hen (1% recombinatie tussen a en b; 3% tussen b en c; 4% tussen a en c). De recombinatiefrequenties zijn ongeveer evenredig (meer…)