Gènes et chromosomes
Les principes classiques de la génétique ont été déduits par Gregor Mendel en 1865, sur la base des résultats d’expériences de sélection sur des pois. Mendel a étudié l’héritage d’un certain nombre de traits bien définis, comme la couleur des graines, et a pu déduire des règles générales pour leur transmission. Dans tous les cas, il a pu interpréter correctement les schémas d’hérédité observés en supposant que chaque caractère est déterminé par une paire de facteurs héréditaires, que l’on appelle aujourd’hui des gènes. Une copie du gène (appelée allèle) spécifiant chaque caractère est héritée de chaque parent. Par exemple, la reproduction de deux souches de pois, l’une ayant des graines jaunes et l’autre des graines vertes, donne les résultats suivants (figure 3.1). Les souches parentales possèdent chacune deux copies identiques du gène spécifiant les graines jaunes (Y) ou vertes (y), respectivement. Les plantes de la descendance sont donc des hybrides, ayant hérité d’un gène pour les graines jaunes (Y) et d’un autre pour les graines vertes (y). Toutes ces plantes de la descendance (la première génération filiative, ou F1,) ont des graines jaunes, on dit donc que le jaune (Y) est dominant et le vert (y) récessif. Le génotype (composition génétique) des pois de la F1 est donc Yy, et leur phénotype (apparence physique) est jaune. Si une descendance F1 est croisée avec une autre, donnant naissance à une descendance F2, les gènes des graines jaunes et vertes ségrègent d’une manière caractéristique telle que le rapport entre les plantes F2 avec des graines jaunes et celles avec des graines vertes est de 3:1.
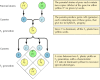
Figure 3.1
Héritage des gènes dominants et récessifs.
Les découvertes de Mendel, apparemment en avance sur leur temps, ont été largement ignorées jusqu’en 1900, lorsque les lois de Mendel ont été redécouvertes et leur importance reconnue. Peu de temps après, le rôle des chromosomes en tant que porteurs de gènes a été proposé. On s’est rendu compte que la plupart des cellules des plantes supérieures et des animaux sont diploïdes – elles contiennent deux copies de chaque chromosome. La formation des cellules germinales (le sperme et l’ovule) implique toutefois un type unique de division cellulaire (méiose) au cours de laquelle un seul membre de chaque paire de chromosomes est transmis à chaque cellule de la descendance (figure 3.2). Par conséquent, le spermatozoïde et l’ovule sont haploïdes, ne contenant qu’une seule copie de chaque chromosome. L’union de ces deux cellules haploïdes lors de la fécondation crée un nouvel organisme diploïde, contenant désormais un membre de chaque paire de chromosomes provenant du parent mâle et un autre du parent femelle. Le comportement des paires de chromosomes est donc parallèle à celui des gènes, ce qui permet de conclure que les gènes sont portés par les chromosomes.
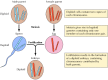
Figure 3.2
Chromosomes à la méiose et à la fécondation. Deux paires de chromosomes d’un organisme hypothétique sont illustrées.
Les principes fondamentaux de la mutation, de la liaison génétique et des relations entre les gènes et les chromosomes ont été largement établis par des expériences réalisées avec la drosophile, Drosophila melanogaster. Les drosophiles peuvent être facilement maintenues en laboratoire, et elles se reproduisent environ toutes les deux semaines, ce qui constitue un avantage considérable pour les expériences génétiques. En effet, ces caractéristiques continuent de faire de la drosophile un organisme de choix pour les études génétiques sur les animaux, en particulier l’analyse génétique du développement et de la différenciation.
Au début des années 1900, un certain nombre d’altérations génétiques (mutations) ont été identifiées chez la drosophile, affectant généralement des caractéristiques facilement observables comme la couleur des yeux ou la forme des ailes. Des expériences d’élevage ont indiqué que certains des gènes régissant ces caractères sont hérités indépendamment les uns des autres, ce qui suggère que ces gènes sont situés sur des chromosomes différents qui se séparent indépendamment au cours de la méiose (figure 3.3). D’autres gènes, en revanche, sont fréquemment hérités ensemble en tant que caractéristiques appariées. On dit de ces gènes qu’ils sont liés entre eux parce qu’ils sont situés sur le même chromosome. Le nombre de groupes de gènes liés est le même que le nombre de chromosomes (quatre chez la drosophile), ce qui appuie l’idée que les chromosomes sont porteurs des gènes.

Figure 3.3
Ségrégation et liaison des gènes. (A) Ségrégation de deux gènes hypothétiques pour la forme (A/a = carré/rond) et la couleur (B/b = rouge/bleu) situés sur des chromosomes différents. (B) Liaison de deux gènes situés sur le même chromosome.
La liaison entre les gènes n’est cependant pas complète ; les chromosomes échangent du matériel pendant la méiose, ce qui entraîne une recombinaison entre les gènes liés (figure 3.4). La fréquence de recombinaison entre deux gènes liés dépend de la distance qui les sépare sur le chromosome ; les gènes proches les uns des autres se recombinent moins fréquemment que les gènes plus éloignés. Ainsi, les fréquences de recombinaison de différents gènes peuvent être utilisées pour déterminer leur position relative sur le chromosome, ce qui permet de construire des cartes génétiques (figure 3.5). En 1915, près d’une centaine de gènes avaient été définis et cartographiés sur les quatre chromosomes de la drosophile, conduisant à l’acceptation générale de la base chromosomique de l’hérédité.
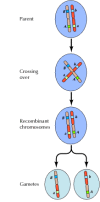
Figure 3.4
Recombinaison génétique. Au cours de la méiose, les membres des paires de chromosomes échangent du matériel. Il en résulte une recombinaison entre les gènes liés.
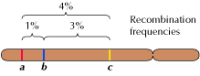
Figure 3.5
Une carte génétique. Trois gènes sont localisés sur un chromosome hypothétique en fonction des fréquences de recombinaison entre eux (1% de recombinaison entre a et b ; 3% entre b et c ; 4% entre a et c). Les fréquences de recombinaison sont approximativement proportionnelles (plus…)
.