Pářící se můry (Tineidae, Ditrysia, Lepidoptera) zachované jako zmrazené chování v baltském jantaru (eocén)
Thilo C. Fischer a Marie K. Hörnig
Číslo článku: 22.1.7
https://doi.org/10.26879/829
Copyright Paleontological Society, únor 2019
Životopisy autorů
Jazykové a vícejazyčné abstrakty
PDF verze
Podání: 22. října 2017. Přijetí: 23. ledna 2019
{flike id=2421}
ABSTRACT
Zkoumání jantarových inkluzí poskytuje jedinečný vhled do biologie živočichů žijících před miliony let, a to díky jejich výjimečnému zachování v téměř životních podmínkách. To se netýká pouze morfologických aspektů, z těchto fosilií lze odvodit i aspekty chování.
Z eocenního baltského jantaru jsou zde uváděny mikrolepidoptery, které jsou s největší pravděpodobností prvním výskytem fosilních Lepidopter nalezených v kopuli. Můry patří do čeledi Tineidae (tineidní můry zahrnují šatní můry) a jsou popsány jako nový druh a nový rod Forcepsites michalskii. Svěrací orgán samců je u můr z baltského jantaru poměrně neobvyklý, má ventrální a dorzální část, z nichž každá je zakončena dvěma paralelními trnovitými výběžky a zřejmě není tvořena valvami. Společný výskyt samce a samice, nejspíše v kopuli, umožnil přímé studium jejich pohlavního dimorfismu a existuje důkaz, že stanovištěm tohoto druhu je místo produkce pryskyřice v baltském jantarovém lese. Z pohlavního dimorfismu tykadel a morfologie očí se interpretuje, že u tohoto druhu byla samice volajícím pohlavím pro páření.
Thilo C. Fischer. Förderverein der Bayerischen Staatssammlung für Paläontologie und Historische Geologie München Richard-Wagner-Straße 10, D-80333 München, Německo.
Marie K. Hörnig. Univerzita Greifswald, Zoologický ústav a muzeum, cytologie a evoluční biologie, Soldmannstr. 23, D-17489 Greifswald, Německo.
Klíčová slova: šatní můry; stanoviště v kopuli; Microlepidoptera; nové druhy a rody; pohlavní dimorfismus
Konečná citace: Fischer, Thilo C, and Hörnig, Marie K. 2019. Pářící se můry (Tineidae, Ditrysia, Lepidoptera) zachované jako inkluze zmrzlého chování v baltském jantaru (eocén). Palaeontologia Electronica 22.1.7A 1-11. https://doi.org/10.26879/829
palaeo-electronica.org/content/2019/2421-mating-moths
Copyright: Únor 2019 Paleontologická společnost.
Jedná se o článek s otevřeným přístupem šířený za podmínek Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), které uživatelům umožňují kopírovat a šířit materiál v jakémkoli médiu nebo formátu za předpokladu, že nebude použit pro komerční účely a bude uveden původní autor a zdroj s uvedením případných změn.
creativecommons.org/licenses/by-nc-sa/4.0/
http://zoobank.org/BCF64D89-6879-4805-8021-7CE926CE4B12
ÚVOD
Pářící se hmyz se v jantaru vzácně vyskytuje jako případy tzv. zamrzlého chování (Arillo, 2007, a odkazy v něm). Takoví byli zaznamenáni z eocenního baltského jantaru, například ze skupin Diptera, Coleoptera, Hemiptera, Hymenoptera a zcela výjimečně Trichoptera (Weitschat a Wichard, 2002; Weitschat, 2009; Boucot a Poinar Jr., 2010; Gröhn, 2016).
V baltském jantaru jsou Lepidoptera obecně vzácní a vyskytují se přibližně v jedné z 250 inkluzí (pro srovnání: Dipterans představují asi 64 % veškerého hmyzu v baltském jantaru, Lepidoptera představují asi 0,4 %; Hoffeins a Hoffeins 2003; Gröhn 2016). Stejně jako u ostatního hmyzu jsou imaga zachovaná v kopuli mnohem vzácnější a Lepidoptera v kopuli dosud nebyla popsána, a to ani v jiných jantarech nebo z jiných než jantarových lokalit hmyzu (Sohn et al., 2012; Sohn a Lamas, 2013). Výskyt in copula umožňuje bona fide příslušnost obou pohlavních forem k sobě navzájem a obou dohromady k samostatnému druhu, a tedy i přesný popis pohlavního dimorfismu druhu. Alternativně lze pohlavní dimorfismus z fosilního hmyzu studovat statistickými metodami pouze vždy, když jsou nálezy daného druhu hojné, jako je tomu u mnoha Nematocera (např, Borkent, 1995).
Tineidae jsou vysoce divergentní bazální zástupci skupiny lepidopter Ditrysia (Regier et al., 2014), jejich larvy se živí širokou škálou substrátů (detrit, houby, lišejníky nebo keratin) (Scoble, 1995), jsou tedy generalisty (85,7 %) (Menken et al., 2009). Jako inkluze baltského jantaru představují Tineidae asi čtvrtinu všech nálezů lepidopter (Skalski, 1977). Zde je popsán vzácný a nový druh Tineidae s jeho pohlavními formami z domnělého výskytu v kopuli.
MATERIÁLY A METODY
Nález jantaru byl nalezen a zakoupen od Artura Michalského (Wroclaw, Polsko). Byl konzervován ošetřením akrylovým lakem a pochází ze sbírky autora (TF) se sbírkovým číslem 7131. Skladování probíhalo při konstantní teplotě v plastových připínacích sáčcích v kovových krabicích s vyloučením přístupu kyslíku a světla. Bude uložen v Bayerische Paläontologische Staatssammlung München pod číslem SNSB-BSPG 2017 I 113.
Vzorek byl vyfotografován fotoaparátem Canon EOS 70d vybaveným objektivem Canon MP-E 65 mm a bleskem Canon Twin Flash MT-24.
Vzorek byl vyfotografován fotoaparátem Canon EOS 70d vybaveným objektivem Canon MP-E 65 mm. Aby se zabránilo odrazům, byly před blesky a objektiv fotoaparátu namontovány a nastaveny polarizační filtry (křížově polarizované světlo). Pro snížení konvexity a konkávnosti jantarového povrchu byla nanesena kapka glycerinu a překryta krycím sklíčkem. Zaznamenané z-stacky snímků byly sloučeny pomocí programu Combine ZP a dále zpracovány v programu Adobe Photoshop CS 3. Inkluzi nebylo možné studovat pomocí mikropočítačové tomografie z důvodu přítomnosti kovových, zřejmě pyritických inkluzí, které způsobují silné artefakty.
SYSTEMATICKÁ PALEONTOLOGIE
Systematika (podle van Nieukerkena et al.), 2011)
Řád LEPIDOPTERA Linnaeus, 1758
Člen DITRYSIA Börner, 1925
Nadčeleď TINEOIDEA Latreille, 1810
Člen TINEIDAE Latreille, 1810
Rod Forcepsites nov. gen.
zoobank.org/DDB6884D-845A-4A87-981E-A2953ADD8964
Etymology. „Forceps“, latinsky kleště, odkazující na samčí svírací orgán.
Druh Forcepsites michalskii nov. spec.
zoobank.org/5494D58E-7773-473C-AFB2-140DDD049226
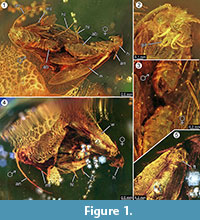 Holotypus. Exemplář ex coll. Fischer č. 7131, obr. 1.2
Holotypus. Exemplář ex coll. Fischer č. 7131, obr. 1.2
Locus typicus. Jantarový důl Jantarni, GUS
Stratum typicum. „Blaue Erde“ (svrchní eocén – spodní oligocén)
Etymologie. Druhové jméno michalskii je zvoleno jako poděkování Arturovi Michalskému za poskytnutí tohoto exempláře.
Repozitář. Bayerische Staatssammlung für Paläontologie und Geologie, Mnichov, Německo; přírůstkové číslo SNSB-BSPG 2017 I 113.
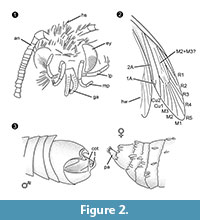
Diagnostika rodu. Monotypický rod s upínacím orgánem samce (obr. 2.3) s ventrální a dorzální částí, z nichž každá je zakončena dvěma paralelními trnovitými výběžky, jako určujícím znakem.
 Diagnostika druhu. Byly určeny příslušné kvalitativní a kvantitativní znaky samičích a samčích imag (obr. 1), které jsou shrnuty v tabulce 1.
Diagnostika druhu. Byly určeny příslušné kvalitativní a kvantitativní znaky samičích a samčích imag (obr. 1), které jsou shrnuty v tabulce 1.
Diferenciální diagnóza. Kombinace znaků malý chobot s oddělenými galejemi, dlouhé a složené maxilární palpy s pěti segmenty, nahoru směřující a mírně ohnuté labiální palpy s laterálními štětinami, hlava s mnoha vzpřímenými chlupovitými šupinami silně naznačují příslušnost k čeledi Tineidae, ale žádný z nich není apomorfní, ani taková apomorfie není v současnosti u dospělých Tineidae známa (Regier et al., 2014). Žilky R4 a R5 nejsou srostlé / stopkaté, což by svědčilo pro Gelechioidea (Gelechiidae a Oecophoridae) (Scoble, 1995), druhou hlavní skupinu Lepidoptera nalezenou v baltském jantaru. Tento druh se nepodobá žádnému popsanému zástupci čeledi Tineidae ani jiných čeledí z baltského jantaru (Rebel, 1934, 1935; Kuzněcov, 1941; Skalski, 1973; Kozlov, 1987, 1988; Sohn et al., 2012; Sohn a Lamas, 2013), zejména pokud jde o samčí pohlavní aparát. Nejpodobnější se zdá být Tineosemopsis decurtatus Skalski 1974 (známá samice), liší se však absencí vzpřímených chlupů na temeni a menšíma očima. Tillyardinea eocaenica Kusnezov 1941 (samice), která má vzpřímené šupiny na temeni a podobnou žilnatinu, se liší frons a labiálními palpy. Simulotinea intermedia Skalski 1977 má podobný habitus, ale liší se také přítomností rozdvojené žilky, která rozděluje velkou buňku v předním křídle (M2+3).
Taphonomy. Samec a samice druhu Forcepsites michalskii jsou fosilizováni blízko sebe a se zadní částí břicha směřující k sobě. Obě inkluze jsou ve válcovitém kusu neautoklávovaného baltského jantaru o průměru 15 mm a výšce 11 mm, který je přibližně v polovině výšky rozdělen nerovným okrajem mezi dvěma vrstvami pryskyřičného toku. Tato hranice vykazuje jako součást své roviny vrstvu pyritu. Jeden pryskyřičný proud obsahuje dvě můry, které vystavují své hřbetní strany této hranici. Zachovány jsou dlouhé šupiny odvozené od okraje samičího křídla. Samičí imago přišlo o dvě nohy a jeho trup je poškozen. Druhý tok obsahuje nelineární řadu bublinek. Oba jedinci se zdají být nově vylíhlí, jak se soudí podle kompletního hávu šupin na křídlech a podle toho, že křídla nejsou potrhaná. Na křídlech, břiše ani nohách nebylo možné identifikovat žádné pachové orgány. Jediné syninkluze zachované u Microlepidoptera jsou trichomy, typické pro baltský jantar.
DISCUSSION
Random Co-occurrence or a Preserved Mating?
Přiložené můry jsou samčí a samičí zástupci téhož druhu a jsou uspořádány blízko sebe s břichy orientovanými k sobě. Přesto nejsou pohlavní orgány můr v přímém kontaktu, ale v těsném postavení. Hřbetní/ventrální orientace samců a samic se mírně liší, což je způsobeno mírnou rotací v ose těla (zejména u samců). Orientace zadní části břicha obou motýlů je směrem k sobě.
Je známo několik případů, kdy situace uzavření v nehybné tekuté pryskyřici může vyvolat stresovou reakci, např. ovipozici (Weitschat, 2009). Vyvolání pářícího chování proudem pryskyřice není obecně pravděpodobné, ale situace uzavření ukazuje, že mírné pohyby v čerstvé pryskyřici byly možné.
Je také otázka, zda toto zjištění může představovat i náhodný souběh. Pravděpodobnost takového náhodného souvýskytu lze přibližně odhadnout. Mezi inkluzemi hmyzu se můry v baltském jantaru vyskytují s frekvencí přibližně jedné z 250 inkluzí (Krzeminskaet al., 1991 (0,6 %); Hoffeins a Hoffeins, 2003 (0,4 %); Sontag, 2003 (0,27 %) a vlastní pozorování). Vzhledem k jednotlivému jantaru se zařazením můry je šance na náhodný společný výskyt další můry jednoduše 1:250 (můry se nevyskytují v rojích). Společný výskyt stejného nebo hojného taxonu, který by mohl být zaměněn, se odhaduje v nejlepším případě na 1:10 (Skalski, 1977) a další asi 1:2 pro výskyt různých pohlaví. Vezmeme-li v úvahu další šance na blízké uspořádání, a to jak zarovnání tělních os, tak vhodnou orientaci těla, je pravděpodobnost náhodného společného výskytu, který by mohl být mylně interpretován jako páření, mnohem nižší než zjištěná frekvence jako jeden z 920 lepidopterních inkluzů ve studované autorské sbírce.
Závěrem lze říci, že nejpravděpodobnější interpretací tohoto nálezu je, že můry byly v kopulaci a genitálie byly v kontaktu, ale jedinci byli od sebe mírně vzdáleni v důsledku toku pryskyřice (viz diskuse níže).
Tafonomie
Význam pečlivé analýzy pryskyřičných toků pro tafonomii jantarových inkluzí prokázali Coty et al. (2014) svou studií interakce mravenec – termit zachované v mexickém jantaru Chiapas.
Výskyt události in copula silně podporuje nález dlouhých šupin odvozených od okraje samičího křídla, které byly na určitou vzdálenost unášeny směrem k samci. To by mohlo svědčit o silném laminárním proudění pryskyřice v tomto místě, které se pohybovalo podél jejich společné osy a mohlo je od sebe odtáhnout na nyní pozorovanou krátkou vzdálenost. Během páření můr může samec ztratit kontakt s místem páření a může být k samici připevněn pouze svěrným orgánem, přičemž pouze samice naváže kontakt s povrchem. Takové chování a přetahování náhlým proudem pryskyřice by mohlo být důvodem, proč samice ztratila obě zadní nohy. Na hřbetní straně můry, která byla obnažena na konvexním povrchu tohoto prvního laminárního proudu pryskyřice, je křídlo samice a pod ním ležící hrudník částečně poškozen (obr. 1.5). Předpokládejme, že tato destruovaná místa představují stopy po žíru způsobené jiným hmyzem, nejspíše mravenci, jak je často pozorováno u exemplářů jantaru. Z toho vyplývá, že po prvním toku pryskyřice jako události usazování budou hřbetní strany po určitou dobu obnaženy, a to dostatečně dlouho na to, aby se po tomto pevném povrchu mohli pohybovat predátoři. Druhý tok pryskyřice, který pokryl tuto stranu později, mohl být rychlým a turbulentním tokem pryskyřice, jak naznačuje řada malých bublinek uspořádaných do nepravidelných křivek, které se v něm nacházejí. To by byl typický tafonomický proces pro takzvanou „Schlaube“ baltského jantaru (Gröhn, 2016).
Sexuální dimorfismus
Sexuální dimorfismus je obtížné pozorovat u fosilních druhů, od nichž je obvykle známo jen několik exemplářů. Znaky neovlivněné pohlavím musí být diagnostické, aby umožnily bezpečné určení druhu jako základ pro další rozlišování pohlaví a objasnění pohlavního dimorfismu. Vzhledem k vysoce pohlavně dimorfním dochovaným druhům, např. s brachypterními samicemi jako u mnoha Psychidae (Scoble, 1995), by byl závěr o příslušnosti obou pohlaví k jednomu druhu sotva možný. Proto lze u fosilních druhů spolehlivě rozpoznat pouze případy slabého pohlavního dimorfismu statistickými prostředky a bez jejich zjištění v kopulaci. To podtrhuje význam zjištěného párovacího chování u fosilních druhů pro popis takových případů silného pohlavního dimorfismu. Takto dochované nálezy fosilního páření mohou také schvalovat korelaci dříve známých pohlavně dimorfních forem k příslušnosti k jednomu druhu, a to i bez možnosti větší statistické podpory. Existuje určitá šance, že dochované páření může představovat mezidruhové nebo mezigenerační páření, jak je známe u můr a motýlů (Ganyard a Brady, 1972; Deering a Scriber, 2002). Tyto jevy jsou však mnohem méně časté než vnitrodruhové páření a sotva je lze nalézt jako vzácné fosilní zařazení.
Nealometrickým pohlavním dimorfismem pozorovaným u Forcepsites michalskii jsou oddělené galeje v chobotu u samců. To může naznačovat nefunkční stav, kdy samčí imága tohoto druhu jsou nežravá, jak je vidět u mnoha existujících (samců) Psychidae s vestigiálním nebo chybějícím chobotem (Scoble, 1995). Alometrické pohlavní dimorfismy, které zde byly zjištěny, jsou: (1) Samice je větší než samec, a to jak ve velikosti těla, tak v délce křídel. Příslušné poměry jsou však téměř stejné. (2) U samice je břicho relativně i absolutně širší ve srovnání se samcem. (3) Tykadla samce jsou mnohem větší a silnější než u imaga samice. U několika žijících druhů jsou tykadla pohlavně dimorfní (Scoble, 1995), dobře vyvinutá samčí tykadla slouží k chemosenzoraci samičích feromonů. (4) Čelistní palpy samců jsou dvakrát silnější než u samic a mají žebernatou strukturu. (5) Oči samců jsou také větší, ale počet ommatidií je přibližně stejný. Větší oči pouze přijímají více světla a mohly by být citlivější; rozlišení by však bylo stejné.
Rozpoznávání volajících samiček probíhá prostřednictvím chemoreceptorů samců, zatímco rozpoznávání na krátkou vzdálenost se místo toho opírá o vizuální identifikaci (Groot, 2014). U druhu Forcepsites michalskii se také zdá, že volajícím pohlavím byla samice – čemuž nasvědčuje větší tykadlový aparát a oči samce.
Habitat
Na základě výše uvedené diskuse je zřejmé, že jedinci zachycení v kopulaci se mohou vyskytovat pouze v místě, kde tento druh žil a pářil se. Jednotliví jedinci mohou být unášeni větrem nebo navštěvovat jiné než stanovištní lokality, jako je tomu u vodního hmyzu, který se v baltském jantaru vyskytuje jen zřídka (Wichardet al., 2009); v případě pářících se jedinců by se to naopak sotva mohlo stát, aniž by se oddělili. Zdá se, že v literatuře nejsou žádné důkazy o páření můr za letu. Naopak u většiny motýlů (Lepidoptera) a u převážně nočních mikrolepidopter (můr) samice „přivolávají“ samce feromony a určují tak místa páření (Groot, 2014). Před pářením mohou létat, ale ve většině případů se zdá, že jsou přisedlé a vějířovitě rozdělují feromony. U některých taxonů však mohou být volající nebo hledající pohlaví i obrácená, ale u čeledi Tineidae se uvádí, že volajícím pohlavím je samice (Wagner a Rosovský, 1991; Scoble, 1995). V každém případě místo toku pryskyřice, tj. biotop se stromy produkujícími jantar, bylo s největší pravděpodobností také stanovištěm zde popsané můry šatní.
ZÁVĚR
A tineoid moth with the remarkable male genital apparatus is described as the new species and genus Forcepsites michalskii from Eocene Baltic amber. Samice a samec jsou zachyceni společně v těsné poloze a s břichem orientovaným k sobě, což naznačuje, že tyto můry byly během páření zachyceny v čerstvé pryskyřici. Silné laminární proudění pryskyřice, které naznačuje několik unášených částí těla samice, je mohlo od sebe odtáhnout na krátkou vzdálenost, kterou nyní pozorujeme. Tento exemplář by tedy mohl představovat první příklad fosilní Lepidoptery zachycené v kopuli.
Předpokládá se, že Forcepsites michalskii byl lesní druh, protože jedince zachycené v kopuli lze doložit pouze z místa, kde tento druh žil a pářil se.
Samci a samci druhu F. michalskii vykazují pohlavní dimorfismus ve velikosti těla, délce křídel, proporcích břicha, velikosti složených očí a morfologii čelistních palpsů i tykadel. Na základě zřetelně větších a silnějších tykadel samce, stejně jako na základě jeho větších očí, se předpokládá, že by u tohoto druhu mohla být volajícím pohlavím samice, jak se uvádí i u existujících druhů čeledi Tineidae.
PŘÍSPĚVKY
A. Michalski (Wroclaw, Polsko) patří poděkování za poskytnutí tohoto výjimečného exempláře a upozornění na něj. M. Schwarz z Oberösterreichisches Landesmuseum (Linec, Rakousko) laskavě podpořil screening jejich sbírky Lepidoptera. MKH bylo laskavě financováno stipendiem Studienstiftung des deutschen Volkes a děkuje za podporu S. Harzschovi (Greifswald, Německo). Děkujeme za cenné vědecké připomínky M. Heikkilä a C. Haug (Mnichov, Německo), které k rukopisu učinili, stejně jako za připomínky J. Kriegera (Greifswald, Německo) a za podporu C. MacGraphery (Freising, Německo) a Y. Goldmana (Collinsville, CT, USA) při kontrole anglického textu jako rodilých mluvčích. Dále děkujeme dvěma anonymním recenzentům za jejich užitečné připomínky.
Arillo, A. 2007. Paleoetologie: zkamenělé chování v jantaru. Geologica Acta, 5(2):159-166. https://doi.org/10.1344/105.000000301
Börner, C. 1925. Lepidoptera, Schmetterlinge, s. 358-387. In Brohmer, P. (ed.). Fauna von Deutschland. Ein Bestimmungsbuch unserer heimischen Tierwelt. Quelle und Meyer, Leipzig.
Borkent, A. 1995. Kousalky v křídovém jantaru Severní Ameriky (Diptera: Ceratopogonidae). Backhuys Publishers, Leiden, Nizozemsko.
Boucot, A.J. a Poinar Jr, G.O. 2010. Kompendium fosilního chování. CRC Press, Boca Raton, s. 147-151.
Coty, D., Aria, C., Garrouste, R., Wils, P., Legendre, F. a Nel, A. 2014. První syninkluze mravenců v jantaru s analýzou tafonomie pomocí CT skenu. PLOS one, 9(8):e104410. https://doi.org/10.1371/journal.pone.0104410
Deering, M.D. a Scriber, J.M. 2002. Terénní biologické testy ukazují asymetrii heterospecifických párovacích preferencí mezi hybridizujícími severoamerickými druhy motýlů rodu Papilio (Lepidoptera: Papilionidae). Journal of Ethology 20:25-33. https://doi.org/10.1007/s10164-002-0050-2
Fletcher, T.B. 1940. New generic names for microlepidoptera [Nová rodová jména pro mikrolepidoptery]. Entomologist’s Record, 52:17-19.
Ganyard, M.C. a Brady, Jr U.E. 1972. Interspecifická přitažlivost u Lepidoptera v terénu. Annals of the Entomological Society of America, 65 (6): 1279-1282. https://doi.org/10.1093/aesa/65.6.1279
Gröhn, C. 2016. Inkluze v baltském jantaru. Wachholtz Verlag – Murmann Publishers, Kiel, Hamburg.
Groot, A.T. 2014. Cirkadiánní rytmy sexuálních aktivit u můr: Recenze. Frontiers in Ecology and Evolution 2:43. https://doi.org/10.3389/fevo.2014.00043
Hoffeins, C. a Hoffeins, H.W. 2003. Studie o množství inkluzí v baltském a bitterfeldském jantaru (třetihory, eocén) z nevybraných sbírek se zvláštním zřetelem k řádu Diptera. Studia Dipterologica, 10(2):381-392.
Kozlov, M.V. 1987. Noví motýlkovití Lepidoptera z baltského jantaru. Paleontological Journal, 4:59-67.
Kozlov, M.V. 1988. Paleontologie lepidopter a problémy fylogeneze řádu Papilionida, s. 16-69. In Ponomarenko A.G. (ed.), The Mesozoic-Cenozoic Crisis in the Evolution of Insects. Akademie věd, Moskva.
Krzeminska, E., Krzeminski, W., Haenni, J.-P., and Dufour, C. 1991. Les Fantomes De L’Ambre. Muséed’histoire naturelle de Neuchâtel, Neuchâtel, Švýcarsko.
Kusnezov, N. 1941. Revize jantarových motýlů. Paleontologický ústav Akademie věd SSSR, Moskva & Leningrad.
Latreille, P.A. 1810. Obecné úvahy o přirozeném uspořádání živočichů tvořících třídy korýšů, pavoukovců a hmyzu: s metodickou tabulkou jejich rodů uspořádaných do čeledí. Paris, Schoell.
Menken, S.B.J., Boomsma, J.J. a van Nieukerken, E.J. 2009. Velkoplošné evoluční vzorce asociací hostitelských rostlin u motýlů (Lepidoptera). Evolution, 64(4):1098-1119. https://doi.org/10.1111/j.1558-5646.2009.00889.x
Rebel, H. 1934. Bernstein-Lepidopteren (aus der Sammlung Bachofen-Echt). Palaeobiologica, 6:1-16.
Rebel, H. 1935. Bernstein-Lepidopteren. Entomologische Zeitschrift Iris, 49:162-186.
Regier, J.C., Mitter, C., Davis, D.R., Harrison, T.L., Sohn J.-C., Cummings, M.P., Zwick, A., and Mitter, K.T. 2014. A molecular phylogeny and revised classification for the oldest ditrysian moth lines (Lepidoptera: Tineoidea), with implications for ancestral feeding habits of the mega-diverse Ditrysia. Systematic Entomology, 40(2). https://doi.org/10.1111/syen.12110
Scoble, M.J. 1995. Lepidoptera – forma, funkce a rozmanitost. The Natural History Museum / Oxford University Press, Oxford.
Skalski, A.W. 1973. Studies on the Lepidoptera from fossil resins [Studie o motýlech z fosilních pryskyřic]. Část II. Epiborkhausenites obscurotrimaculatus gen. et sp. nov. (Oecophoridae) and a Tineid-moth discovered in Baltic amber. Acta Palaeontologica Polonica, 18(1):153-160.
Skalski, A.W. 1974. Zwei neue Gattungen und Arten der Familie Tineidae aus dem Baltischen Bernstein. Studien an Lepidopteren aus fossilen Harzen V. Beiträge zur Entomologie, 1/4(24):97-104.
Skalski, A.W. 1977. Studie o motýlech (Lepidoptera) z fosilních pryskyřic. Part I. General remarks and descriptions of new genera and species of the families Tineidae and Oecophoridae from Baltic amber. Prace Museum Ziemi, 26:3-24.
Sohn, J.-C., Labandeira, C., Davis, D., and Mitter, C. 2012. An annotated catalog of fossil and subfossil Lepidoptera (Insecta: Holometabola) of the world (Komentovaný katalog fosilních a subfosilních motýlů (Lepidoptera) světa). Zootaxa, 3286:1-132.
Sohn, J.-C. a Lamas, G. 2013. Opravy, doplňky a nomenklatorické poznámky k nedávno vydanému světovému katalogu fosilních a subfosilních motýlů (Lepidoptera). Zootaxa, 3599:395-399.
Sontag, E. 2003. Živočišné inkluze ve vzorku nevybraného baltského jantaru. Acta Zoologica Cracoviensia, 46 (suppl.-Fossil Insects):431-440.
van Nieukerken, E.J., Kaila, L., Kitching, I.J., Kristensen, N.P., Lees, D.C., Minet, J., Mitter, C., Mutanen, M., Regier, J.C., Simonsen, T.J., Wahlberg, N., Yen, S.-H., Zahiri, R., Adamski, D., Baixeras, J., Bartsch, D., Bengtsson, B.Å., Brown, J.W., Bucheli, S.R., Davis, D.R., De Prins, J., De Prins, W., Epstein, M.E., Gentili-Poole, P., Gielis, C., Hätteschwiler, P., Hausmann, A., Holloway, J.D., Kallies, A., Karsholt, O., Kawahara, A.Y., Koster, S.J.C., Kozlov, M.V., Lafontaine, J.D., Lamas, G., Landry, J.-F., Lee, S., Nuss, M., Park, K.-T., Penz, C., Rota, J., Schintlmeister, A., Schmidt, B.C., Sohn, J.-C, Solis, M.A., Tarmann, G.M., Warren, A.D., Weller, S., Yakovlev, R.V., Zolotuhin, V.V., and Zwick, A. 2011. Order Lepidoptera Linnaeus, 1758. In Zhang, Z.-Q. (ed.), Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa, 3148:212-221.
von Linné, C. 1758. překlad William Turton (1806). Svazek 1. Obecný systém přírody: Tři velké říše živočichů, rostlin a nerostů, systematicky rozdělené do několika tříd, řádů, rodů, druhů a odrůd. Lackington, Allen, and Co, London.
Wagner, D.L. a Rosovsky, J. 1991. Mating systems in primitive Lepidoptera, with emphasis on the reproductive behaviour of Korscheltellus gracilis (Hepialidae) [Systémy páření u primitivních motýlů (Lepidoptera) s důrazem na reprodukční chování Korscheltellus gracilis (Hepialidae)]. Zoological Journal of the Linnean Society, 102:277-303. https://doi.org/10.1111/j.1096-3642.1991.tb00003.x
Weitschat, W. a Wichard, W. 2002. Atlas rostlin a živočichů v baltském jantaru. Verlag Friedrich Pfeil, München.
Weitschat, W. 2009. Jäger, Gejagte, Parasiten und blinde Passagiere – Momentaufnahmen aus dem Bernsteinwald. Denisia, 26(86):243-256.
Wichard, W., Gröhn, C. a Seredszus, F. 2009. Vodní hmyz v baltském jantaru. Verlag Kessel, Remagen-Oberwinter.
.