- Pariutumisperhoset (Tineidae, Ditrysia, Lepidoptera), jotka ovat säilyneet jäätyneinä käyttäytymissulkeumina Baltian meripihkassa (eoseeni)
- ABSTRACT
- INTRODUCTION
- MATERIAALIT JA MENETELMÄT
- SYSTEMATIC PALAEONTOLOGY
- KESKUSTELU
- Sattumanvarainen yhteisesiintyminen vai säilynyt pariutuminen?
- Tafonomia
- Sukupuolidimorfismi
- Habitaatti
- YHTEENVETO
- TAKAISINKIRJOITUKSET
Pariutumisperhoset (Tineidae, Ditrysia, Lepidoptera), jotka ovat säilyneet jäätyneinä käyttäytymissulkeumina Baltian meripihkassa (eoseeni)
Thilo C. Fischer ja Marie K. Hörnig
Artikkelinumero: 22.1.7
https://doi.org/10.26879/829
Copyright Paleontological Society, February 2019
Author biographies
Plain-language- and multi-lingual abstracts
PDF-versio
Submission: 22 Octobere 2017. Acceptance: 23 Januiary 2019
{flike id=2421}
ABSTRACT
Mustikkasulkeumien tutkiminen tarjoaa ainutlaatuisen näkemyksen miljoonia vuosia sitten eläneiden eläinten biologiaan, koska ne säilyvät poikkeuksellisen hyvin lähes elämän kaltaisissa olosuhteissa. Tämä ei koske ainoastaan morfologisia näkökohtia, vaan myös käyttäytymiseen liittyviä näkökohtia voidaan päätellä näistä fossiileista.
Tässä raportoidaan Eoseenin balttilaisesta meripihkasta löydetyt Microlepidopterat, jotka ovat mitä todennäköisimmin ensimmäinen kopulasta löydetty fossiilisten Lepidopteroiden esiintymä. Perhoset kuuluvat Tineidae-heimoon (tineid-perhosiin kuuluvat mm. vaateperhoset) ja ne kuvataan uutena lajina ja uutena sukuna, Forcepsites michalskii. Uroksen kiinnityselin on varsin epätavallinen Itämeren meripihkakammioperhosille, sillä siinä on ventraalinen ja dorsaalinen osa, jotka kumpikin päättyvät kahteen samansuuntaiseen, piikkimäiseen ulokkeeseen, ja se ei näytä muodostuvan valvaeista. Uroksen ja naaraan samanaikainen esiintyminen, todennäköisesti kopulassa, mahdollisti niiden sukupuolidimorfismin suoran tutkimisen, ja on viitteitä siitä, että tämän lajin elinympäristö on hartsin tuotantopaikka Baltian meripihkametsissä. Antennien ja silmien morfologian sukupuolidimorfismin perusteella tulkitaan, että naaras oli tässä lajissa kutsuva sukupuoli parittelua varten.
Thilo C. Fischer. Förderverein der Bayerischen Staatssammlung für Paläontologie und Historische Geologie München Richard-Wagner-Straße 10, D-80333 München, Saksa.
Marie K. Hörnig. Greifswaldin yliopisto, eläintieteellinen instituutti ja museo, sytologia ja evoluutiobiologia, Soldmannstr. 23, D-17489 Greifswald, Saksa.
LYHENTEET: Vaatekotilot; elinympäristö kopulassa; Microlepidoptera; uudet lajit ja suvut; sukupuolidimorfismi
Loppusitaatti: Fischer, Thilo C ja Hörnig, Marie K. 2019. Itämeren meripihkassa (Eoseeni) pakastekäyttäytymisen sulkeutumisena säilyneet parittelevat koiperhoset (Tineidae, Ditrysia, Lepidoptera). Palaeontologia Electronica 22.1.7A 1-11. https://doi.org/10.26879/829
palaeo-electronica.org/content/2019/2421-mating-moths
Copyright: Helmikuu 2019 Paleontologinen seura.
Tämä on open access -artikkeli, jota jaetaan Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0) -lisenssillä, joka sallii käyttäjien kopioida ja levittää aineistoa missä tahansa välineessä tai muodossa edellyttäen, että sitä ei käytetä kaupallisiin tarkoituksiin ja että alkuperäistä kirjoittajaa ja alkuperälähdettä mainitaan sekä ilmoitetaan, jos siihen on tehty muutoksia.
creativecommons.org/licenses/by-nc-sa/4.0/
http://zoobank.org/BCF64D89-6879-4805-8021-7CE926CE4B12
INTRODUCTION
Parittelevia hyönteisiä tavataan meripihkasta harvoin niin sanotun ”jähmettyneen käyttäytymisen” tapauksina (Arillo, 2007, ja viittaukset siihen). Tällaisia on raportoitu eoseenisestä balttilaisesta meripihkasta esimerkiksi ryhmistä Diptera, Coleoptera, Hemiptera, Hymenoptera ja melko harvoin Trichoptera (Weitschat ja Wichard, 2002; Weitschat, 2009; Boucot ja Poinar Jr., 2010; Gröhn, 2016).
Balttilaisesta meripihkasta Lepidopterat ovat yleisesti ottaen harvinaisia, ja niitä esiintyy noin yksi 250 sulkeutumasta (vertailun vuoksi: Dipterans edustavat noin 64 % kaikista Baltian meripihkan hyönteisistä, Lepidoptera noin 0,4 %; Hoffeins ja Hoffeins 2003; Gröhn 2016). Kuten muillakin hyönteisillä, kopulassa säilyneet imagoset ovat paljon harvinaisempia, eikä kopulassa esiintyviä leppäkerttuja ole vielä kuvattu, ei edes muista meripihkoista tai muilta kuin meripihkan hyönteislöytöpaikoilta (Sohn et al., 2012; Sohn ja Lamas, 2013). In copula -esiintyminen mahdollistaa molempien sukupuolimuotojen bona fide liittämisen toisiinsa ja molempien yhdessä erilliseen lajiin ja siten lajin sukupuolidimorfismin tarkan kuvaamisen. Vaihtoehtoisesti fossiilisten hyönteisten sukupuolidimorfismia voidaan tutkia vain tilastollisin keinoin aina, kun löydöksiä lajista on runsaasti, kuten monilla Nematoceroilla (esim, Borkent, 1995).
Tineidae ovat lepidopteraryhmän Ditrysia (Regier ym., 2014) erittäin divergenttejä basaalisia edustajia, joiden toukat syövät monenlaisia substraatteja (detritus, sienet, jäkälät tai keratiini) (Scoble, 1995) ja ovat siten generalisteja (85,7 %) (Menken ym., 2009). Itämeren meripihkan sulkeumina Tineidae edustaa noin neljäsosaa kaikista lepidopteralöydöistä (Skalski, 1977). Tässä kuvataan harvinainen ja uusi Tineidae-laji sukupuolimuotoineen oletetusta in copula -esiintymästä.
MATERIAALIT JA MENETELMÄT
Marmoripihkanäytteen löysi ja osti Artur Michalski (Wroclaw, Puola). Se konservoitiin käsittelemällä se akryylilakalla, ja se on tekijän (TF) kokoelmasta, jonka kokoelmanumero on 7131. Säilytys tapahtui vakiolämpötilassa muovisissa klipsipusseissa metallilaatikoissa ilman happea ja valoa. Se talletetaan Bayerische Paläontologische Staatssammlung Müncheniin numerolla SNSB-BSPG 2017 I 113.
Näyte kuvattiin Canon EOS 70d -kameralla, jossa oli Canon MP-E 65 mm:n objektiivi ja Canon Twin Flash MT-24. Heijastusten välttämiseksi salaman ja kameran objektiivin eteen asennettiin ja säädettiin polarisaatiosuodattimet (ristipolarisoitu valo). Meripihkan pinnan kuperuuden ja koveruuden vähentämiseksi siihen levitettiin tippa glyseriiniä ja se peitettiin peitelevyllä. Tallennetut kuvan z-pinot fuusioitiin Combine ZP:llä ja käsiteltiin edelleen Adobe Photoshop CS 3 -ohjelmalla. Sulkeumaa ei voitu tutkia mikrotietokonetomografialla, koska siinä oli metallisia, ilmeisesti pyriittisiä sulkeumia, jotka aiheuttavat voimakkaita artefakteja.
SYSTEMATIC PALAEONTOLOGY
Systematiikka (van Nieukerken ym. mukaan, 2011)
Luokka LEPIDOPTERA Linnaeus, 1758
Luokka DITRYSIA Börner, 1925
Yläluokka TINEOIDEA Latreille, 1810
Suku TINEIDAE Latreille, 1810
Suku Forcepsites nov. gen.
zoobank.org/DDB6884D-845A-4A87-981E-A2953ADD8964
Etymology. ”Forcepsites”, latinan koura, joka viittaa uroksen puristuselimeen.
Laji Forcepsites michalskii nov. spec.
zoobank.org/5494D58E-7773-473C-AFB2-140DDD049226
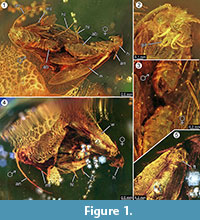 Holotypus. Specimen ex coll. Fischer no. 7131, kuva 1.2
Holotypus. Specimen ex coll. Fischer no. 7131, kuva 1.2
Locus typicus. Yantarnin meripihkakaivos, GUS
Stratum typicum. ”Blaue Erde” (ylempi eoseeni – alempi oligoseeni)
Etymologia. Lajinimi michalskii on valittu kiitokseksi Artur Michalskille tämän yksilön toimittamisesta.
Repositio. Bayerische Staatssammlung für Paläontologie und Geologie, München, Saksa; liittymisnumero SNSB-BSPG 2017 I 113.
Suvun määritys. Monotyyppinen suku, jonka määrittävänä tuntomerkkinä on uroksen tarttumiselin (kuva 2.3), jossa on ventraalinen ja dorsaalinen osa, jotka kumpikin päättyvät kahteen samansuuntaiseen, piikkimäiseen ulokkeeseen.
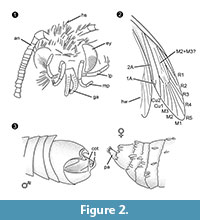 Lajin määritys. Naaras- ja uroskuvioiden (kuva 1) vastaavat laadulliset ja määrälliset tuntomerkit määritettiin, ja niistä on yhteenveto taulukossa 1.
Lajin määritys. Naaras- ja uroskuvioiden (kuva 1) vastaavat laadulliset ja määrälliset tuntomerkit määritettiin, ja niistä on yhteenveto taulukossa 1.
Differentiaalidiagnoosi. Merkkien yhdistelmä pieni kitalaki, jossa on erilliset galeat, pitkät ja taittuneet maksimipalpat, joissa on viisi segmenttiä, ylöspäin suuntautuvat ja hieman taivutetut labiaalipalpat, joissa on sivuharjakset, pää, jossa on paljon pystykarvoja muistuttavia suomuja, viittaavat vahvasti kuulumiseen Tineidae-heimoon, mutta mikään näistä ei ole apomorfinen, eikä tällaista apomorfiaa tunneta tällä hetkellä myöskään aikuisilta tuntemattomilta Tineidae-heimon lajeilta (Regier ym., 2014). Suonet R4 ja R5 eivät ole fuusioituneet/varretut, mikä viittaisi Gelechioideaan (Gelechiidae ja Oecophoridae) (Scoble, 1995), joka on toinen Baltian meripihkasta löydetty Lepidoptera-suvun pääryhmä. Laji poikkeaa erityisesti urospuolisten sukupuolielintensä osalta kaikista Itämeren meripihkasta kuvatuista Tineidae-heimon tai muiden sukujen edustajista (Rebel, 1934, 1935; Kuznezov, 1941; Skalski, 1973; Kozlov, 1987, 1988; Sohn ym., 2012; Sohn ja Lamas, 2013). Kaikkein samankaltaisimmalta näyttää Tineosemopsis decurtatus Skalski 1974 (naaras tiedossa), mutta eroaa siitä pystyjen karvojen puuttumisessa kärkeen ja pienemmissä silmissä. Tillyardinea eocaenica Kusnezov 1941 (naaras), jolla on pystykasvuisia suomuja kärkipisteessä ja samanlainen suonitus, eroaa siitä fronssin ja labiaalisten palpoiden osalta. Simulotinea intermedia Skalski 1977 on habituseltaan samanlainen, mutta eroaa myös siinä, että sillä on haarautuva suoni, joka jakaa etusiiven suuren solun (M2+3).
Tafonomia. Forcepsites michalskii -lajin uros ja naaras ovat fossiloituneet lähelle toisiaan ja vatsan takaosa on suunnattu toisiinsa nähden. Molemmat sulkeumat ovat sylinterimäisessä, halkaisijaltaan 15 mm:n ja korkeudeltaan 11 mm:n läpimittaisessa, autoklavoimattomasta balttilaisesta meripihkasta valmistetussa kappaleessa, jota noin puolet sen korkeudesta jakaa kahden hartsivirtauskerroksen välinen epätasainen raja. Tässä reunassa on pyriittikerros osana sen tasoa. Yhdessä hartsivirtauksessa on kaksi koiperhosta, jotka paljastavat selkäpuolensa tälle rajalle. Naaraan siiven reunasta peräisin olevat pitkät suomut ovat säilyneet. Naaraan imago on menettänyt kaksi jalkaa, ja sen rintakehä on vaurioitunut. Toinen virtaus sisältää epälineaarisen sarjan kuplia. Molemmat yksilöt vaikuttavat vastikään syntyneiltä, mikä voidaan päätellä siipien täydellisestä suomupeitteestä ja siitä, että siivet eivät ole riekaleina. Siivissä, vatsassa tai jaloissa ei havaittu hajuelimiä. Ainoat Microlepidopteran kanssa säilyneet synkliinit ovat trikomeja, jotka ovat tyypillisiä balttilaiselle meripihkalle.
KESKUSTELU
Sattumanvarainen yhteisesiintyminen vai säilynyt pariutuminen?
Ohessa olevat koiperhoset ovat saman lajin uros- ja naaraspuolisia edustajia, ja ne ovat järjestäytyneet lähekkäin siten, että niiden vatsat ovat suuntautuneet toisiaan kohti. Koitten sukupuolielimet eivät kuitenkaan ole suorassa kosketuksessa, vaan lähekkäin. Uroksen ja naaraan dorsaalinen/ventraalinen suuntautuminen eroaa hieman toisistaan, mikä johtuu lievästä kiertymisestä vartalon akselin sisällä (erityisesti uroksen). Molempien perhosten vatsan takaosan suunta on toisiaan kohti.
Tiedetään useita tapauksia, joissa vielä nestemäiseen hartsiin sulkeminen voi aiheuttaa stressireaktion, kuten muninnan (Weitschat, 2009). Parittelukäyttäytymisen indusoiminen hartsivirtauksen vaikutuksesta ei yleensä ole uskottavaa, mutta kotelointitilanne osoittaa, että vähäiset liikkeet tuoreen hartsin sisällä olivat mahdollisia.
Kysymyksenä on myös, voisiko tämä havainto edustaa myös satunnaista yhteisesiintymistä. Tällaisen satunnaisen yhteisesiintymän todennäköisyys voidaan karkeasti arvioida. Hyönteissulkeumien joukossa koiperhosia esiintyy noin yksi 250:stä Baltian meripihkan sulkeumasta (Krzeminskaet al., 1991 (0,6 %); Hoffeins ja Hoffeins, 2003 (0,4 %); Sontag, 2003 (0,27 %) ja oma havainto). Kun kyseessä on yksittäinen meripihka, jossa on koiperhonen, toisen koiperhosen satunnaisen esiintymisen mahdollisuus on yksinkertaisesti 1:250 (koiperhoset eivät esiinny parvissa). Saman taksonin tai erehdyttävästi runsaan taksonin samanaikaisen esiintymisen todennäköisyydeksi arvioidaan parhaimmillaan 1:10 (Skalski, 1977) ja eri sukupuolten esiintymisen todennäköisyydeksi noin 1:2. Kun otetaan huomioon myös mahdollisuudet läheiseen järjestäytymiseen, sekä vartalon akselien suuntautumiseen että vartalon sopivaan suuntautumiseen, todennäköisyys satunnaiselle yhteisesiintymiselle, joka voitaisiin tulkita väärin parittelutapahtumaksi, on paljon pienempi kuin havaittu esiintymistiheys, joka on yksi 920:stä tutkittavan tekijän kokoelman lepidoptera-sulkeumasta.
Johtopäätöksenä todettakoon, että todennäköisin tulkinta tälle havainnolle on se, että koiperhoset olivat kopulaatiossa ja sukupuolielimet olivat olleet kosketuksissa, mutta yksilöt olivat hieman etääntyneet toisistaan hartsivirtauksen vuoksi (ks. keskustelu jäljempänä).
Tafonomia
Hartsivirtojen huolellisen analyysin merkitys meripihkan sulkeumien taphonomian kannalta on osoitettu Coty ym. (2014) tutkimuksessa, joka koski Meksikon Chiapasin meripihkassa säilynyttä muurahaisen ja termiitin vuorovaikutusta.
Kopula-tapahtuman esiintymistä tukee vahvasti naaraspuolisen koiraan siiven reunasta peräisin olevien pitkien hilseiden löytyminen, jotka ajelehtivat jonkun matkan verran urospuolisen koiraan suuntaan. Tämä voisi viitata voimakkaaseen laminaariseen hartsivirtaukseen tässä paikassa, joka liikkui niiden yhteistä akselia pitkin ja olisi voinut vetää ne erilleen nyt havaitun lyhyen matkan verran. Perhosten parittelun aikana uros voi menettää kosketuksen parittelupaikkaan, ja se voi olla naaraaseen kiinnittyneenä vain kiinnityselimen avulla, jolloin vain naaras on kosketuksissa pinnan kanssa. Tällainen käyttäytyminen ja äkillisen hartsivirtauksen aiheuttama vetovoima voivat olla syynä siihen, että naaras on menettänyt molemmat takajalkansa. Naaraan siipi ja sen alla oleva rintakehä ovat osittain vaurioituneet naaraan selkäpuolella, joka oli paljastunut tämän ensimmäisen laminaarisen hartsivirtauksen kuperalla pinnalla (kuva 1.5). Oletettavasti nämä tuhoutuneet alueet ovat muiden hyönteisten, todennäköisesti muurahaisten, aiheuttamia ruokailujälkiä, kuten meripihkanäytteissä usein havaitaan. Ensimmäisen hartsivirtauksen jälkeen selkäpuolet ovat siis olleet jonkin aikaa alttiina, tarpeeksi kauan, jotta saalistajat ovat voineet kävellä tällä kiinteällä pinnalla. Toinen hartsivirtaus, joka peitti tämän puolen myöhemmin, on voinut olla nopea ja turbulenttinen hartsivirtaus, kuten sen sisällä havaitut epäsäännöllisen kaarevat pienet kuplat osoittavat. Tämä olisi tyypillinen taphonominen prosessi Baltian meripihkan niin sanotulle ”Schlaubelle” (Gröhn, 2016).
Sukupuolidimorfismi
Sukupuolidimorfismia on vaikea havaita fossiilisissa lajeissa, joista tunnetaan yleensä vain muutamia yksilöitä. Merkkien, joihin sukupuoli ei vaikuta, on oltava diagnostisia, jotta ne mahdollistavat varman lajimäärityksen, joka on perusta sukupuolten tarkemmalle erottelulle ja seksuaalidimorfismin selvittämiselle. Kun otetaan huomioon erittäin sukupuolisesti dimorfiset nykyiset lajit, esimerkiksi lajit, joissa naaraat ovat brachypteroottisia, kuten monissa Psychidae-heimoissa (Scoble, 1995), olisi tuskin mahdollista päätellä, että molemmat sukupuolet kuuluisivat yhteen lajiin. Näin ollen fossiilisista lajeista voidaan luotettavasti tunnistaa tilastollisin keinoin ja löytämättä niitä kopuloista vain heikon sukupuolidimorfismin tapauksia. Tämä korostaa fossiilisten lajien tunnistetun pariutumiskäyttäytymisen merkitystä tällaisten vahvan seksuaalidimorfismin tapausten kuvaamisessa. Tällaiset säilyneet havainnot fossiilisesta parittelusta voivat myös hyväksyä aiemmin tunnettujen sukupuolisesti dimorfisten muotojen korrelaation kuuluvan yhteen lajiin, vaikka tilastollinen tuki ei olisi kovin suuri. On olemassa jonkinlainen mahdollisuus, että säilynyt parittelutapahtuma voisi edustaa lajien tai sukupolvien välistä parittelua, kuten perhosista ja perhosista tiedetään (Ganyard ja Brady, 1972; Deering ja Scriber, 2002). Nämä ilmiöt ovat kuitenkin paljon harvinaisempia kuin lajinsisäinen pariutuminen, ja on tuskin todennäköistä, että niitä löydettäisiin harvinaisena fossiilisena esiintymänä.
Forcepsites michalskii -lajista havaittu ei-allometrinen sukupuolidimorfismi on uroksen proboscissin erilliset kalvot. Tämä saattaa viitata ei-toiminnalliseen tilaan, jolloin tämän lajin urospuoliset imagosit eivät ole ruokailevia, kuten on nähty monissa olemassa olevissa (urospuolisissa) Psychidae-heimoissa, joilla on vestigiaalinen tai puuttuva proboscis (Scoble, 1995). Tässä havaitut allometriset sukupuolidimorfismit ovat: (1) Naaras on urosta suurempi sekä ruumiin koon että siipien pituuden suhteen. Vastaavat suhteet ovat kuitenkin lähes samat. (2) Naaraan vatsa on suhteellisesti ja absoluuttisesti leveämpi urokseen verrattuna. (3) Uroksen antennit ovat paljon suuremmat ja paksummat kuin naaraan imago. Useilla olemassa olevilla lajeilla antennit ovat sukupuolidimorfisia (Scoble, 1995), hyvin kehittyneet uroksen antennit palvelevat naaraan feromonien kemosensorointia. (4) Uroksen leukahampaat ovat kaksi kertaa paksummat kuin naaraan, ja niissä on uurrettu rakenne. (5) Uroksen silmät ovat myös suuremmat, mutta ommatidioiden määrä on suunnilleen sama. Suuremmat silmät vain vastaanottavat enemmän valoa ja voivat olla herkemmät; resoluutio olisi kuitenkin sama.
Kutsuvien naarasperhosten pitkän kantaman tunnistaminen tapahtuu uroksen kemoreseptorien kautta, kun taas lyhyen kantaman tunnistaminen perustuu sen sijaan visuaaliseen tunnistamiseen (Groot, 2014). Forcepsites michalskii -lajilla näyttää myös siltä, että naaras oli kutsuva sukupuoli – mihin viittaavat urosta suuremmat antennilaitteet ja silmät.
Habitaatti
Yllä esitetyn keskustelun perusteella on selvää, että kopulasta pyydystetyt yksilöt voivat esiintyä vain paikassa, jossa tämä laji asui ja pariutui. Yksittäiset yksilöt voivat ajautua tuulen mukana tai käydä elinympäristön ulkopuolisilla paikoilla, kuten Itämeren meripihkasta harvoin löytyvät vesihyönteiset (Wichardet al., 2009); parittelevien yksilöiden kohdalla tämä sen sijaan tuskin voisi tapahtua ilman erilleen joutumista. Kirjallisuudessa ei näytä olevan todisteita lentävien koiperhosten parittelusta. Sen sijaan useimmilla lepikkoperhosilla ja pääasiassa yöllä elävillä mikrolepidopteroilla (perhosilla) naaraat ”kutsuvat” uroksia feromonien avulla ja määräävät näin parittelupaikat (Groot, 2014). Ne saattavat lentää ennen parittelua, mutta useimmissa tapauksissa ne näyttävät olevan istuvia ja haahuilevia feromonien levittämiseksi. Joissakin taksoneissa kutsuva tai etsivä sukupuoli voi kuitenkin olla myös käänteinen, mutta Tineidae-heimon osalta naaraan on raportoitu olevan kutsuva sukupuoli (Wagner ja Rosovsky, 1991; Scoble, 1995). Joka tapauksessa pihkan virtauspaikka eli biotooppi, jossa oli meripihkaa tuottavia puita, oli todennäköisesti myös tässä kuvatun vaatekoiraan elinympäristö.
YHTEENVETO
Kuvaamme tineoidisen koiraan, jolla on huomattava uroksen sukuelinlaite, uuden lajin ja suvun Forcepsites michalskii eoseenisestä balttilaisesta meripihkasta. Naaras ja uros ovat säilyneet yhdessä lähekkäin ja vatsa toisiinsa päin suuntautuneina, mikä viittaa siihen, että nämä koiperhoset jäivät parittelun aikana kiinni tuoreeseen hartsiin. Voimakas laminaarinen hartsivirtaus, johon viittaavat useat naaraan vartalon ajautuneet osat, olisi voinut vetää ne erilleen nyt havaitun lyhyen matkan päähän. Tämä yksilö voisi siis edustaa ensimmäistä esimerkkiä kopulasta pyydystetystä fossiilisesta lepikopterasta.
On oletettavaa, että Forcepsites michalskii oli metsäeläinlaji, sillä kopulasta pyydystettyjä yksilöitä voidaan dokumentoida vain paikasta, jossa tämä laji eli ja pariutui.
F. michalskii -naaras- ja urospuolisilla lajeilla on sukupuolidimorfismia ruumiin koossa, siipien pituudessa, vatsan mittasuhteissa, yhdistelmäsilmien koossa ja leukapalpettien sekä antennien morfologiassa. Uroksen selvästi suurempien ja paksumpien antennien sekä isompien silmien perusteella ehdotetaan, että naaras voisi olla kutsuva sukupuoli tässä lajissa, kuten on raportoitu myös nykyisistä Tineidae-heimon lajeista.
TAKAISINKIRJOITUKSET
A. Michalski (Wroclaw, Puola) saa kiitokset tämän poikkeuksellisen yksilön toimittamisesta ja huomiomme kiinnittämisestä siihen. M. Schwarz Oberösterreichisches Landesmuseumista (Linz, Itävalta) antoi ystävällisesti tukensa Lepidoptera-kokoelmansa seulontaan. MKH sai ystävällisesti Studienstiftung des deutschen Volkesin stipendin ja kiittää S. Harzschia (Greifswald, Saksa) tuesta. Kiitämme M. Heikkilää ja C. Haugia (München, Saksa) käsikirjoituksesta antamistamme arvokkaista tieteellisistä kommenteista sekä J. Kriegerin (Greifswald, Saksa) kommenteista ja C. MacGrapheryn (Freising, Saksa) ja Y. Goldmanin (Collinsville, CT, Yhdysvallat) äidinkielenään äidinkielenään puhujina antamasta tuesta englanninkielisen tekstin tarkistamisessa. Lisäksi kiitämme kahta anonyymiä arvioijaa hyödyllisistä kommenteista.
Arillo, A. 2007. Paleoetologia: fossiilista käyttäytymistä meripihkassa. Geologica Acta, 5(2):159-166. https://doi.org/10.1344/105.000000301
Börner, C. 1925. Lepidoptera, Schmetterlinge, s. 358-387. Teoksessa Brohmer, P. (toim.). Fauna von Deutschland. Ein Bestimmungsbuch unserer heimischen Tierwelt. Quelle und Meyer, Leipzig.
Borkent, A. 1995. Purevat käävät Pohjois-Amerikan kalkkikautisessa meripihkassa (Diptera: Ceratopogonidae). Backhuys Publishers, Leiden, Alankomaat.
Boucot, A.J. ja Poinar Jr, G.O. 2010. Fossil Behavior Compendium. CRC Press, Boca Raton, s. 147-151.
Coty, D., Aria, C., Garrouste, R., Wils, P., Legendre, F. ja Nel, A. 2014. Ensimmäinen muurahaistermiittisynklima meripihkasta CT-skannausanalyysin taphonomian avulla. PLOS one, 9(8):e104410. https://doi.org/10.1371/journal.pone.0104410
Deering, M.D. ja Scriber, J.M. 2002. Kenttäbiomääritykset osoittavat heterospesifistä parittelupreferenssin epäsymmetriaa risteytyvien pohjoisamerikkalaisten Papilio-perhoslajien (Lepidoptera: Papilionidae) välillä. Journal of Ethology 20:25-33. https://doi.org/10.1007/s10164-002-0050-2
Fletcher, T.B. 1940. Uusia geneerisiä nimiä mikrolepidopteroille. Entomologist’s Record, 52:17-19.
Ganyard, M.C. ja Brady, Jr.U.E. 1972. Interspecific attraction in Lepidoptera in the field. Annals of the Entomological Society of America, 65 (6): 1279-1282. https://doi.org/10.1093/aesa/65.6.1279
Gröhn, C. 2016. Itämeren meripihkan sulkeumat. Wachholtz Verlag – Murmann Publishers, Kiel, Hamburg.
Groot, A.T. 2014. Yöperhosten seksuaalisen toiminnan vuorokausirytmi: Katsaus. Frontiers in Ecology and Evolution 2:43. https://doi.org/10.3389/fevo.2014.00043
Hoffeins, C. ja Hoffeins, H.W. 2003. Itämeren ja Bitterfeldin meripihkan (tertiääri, eoseeni) valikoimattomista kokoelmista peräisin olevien sulkeumien runsautta koskevat tutkimukset, joissa kiinnitetään erityistä huomiota Diptera-luokkaan. Studia Dipterologica, 10(2):381-392.
Kozlov, M.V. 1987. Uusia yöperhosia Itämeren meripihkasta. Paleontological Journal, 4:59-67.
Kozlov, M.V. 1988. Paleontology of lepidopterans and problems of the phylogeny of the order Papilionida, s. 16-69. Teoksessa Ponomarenko A.G. (toim.), The Mesozoic-Cenozoic Crisis in the Evolution of Insects. Tiedeakatemia, Moskova.
Krzeminska, E., Krzeminski, W., Haenni, J.-P., and Dufour, C. 1991. Les Fantomes De L’Ambre. Muséed’histoire naturelle de Neuchâtel, Neuchâtel, Sveitsi.
Kusnezov, N. 1941. A Revision of Amber Lepidoptera. Paleontologian instituutti, Neuvostoliiton tiedeakatemia, Moskova & Leningrad.
Latreille, P.A. 1810. Yleisiä näkökohtia äyriäisten, arachnidien ja hyönteisten luokkiin kuuluvien eläinten luonnollisesta järjestyksestä: sekä metodinen taulukko niiden suvuista, jotka on järjestetty sukuihin. Paris, Schoell.
Menken, S.B.J., Boomsma, J.J. ja van Nieukerken, E.J. 2009. Isäntäkasviyhdistysten laajamittaiset evoluutiomallit lepakkoperhosilla. Evolution, 64(4):1098-1119. https://doi.org/10.1111/j.1558-5646.2009.00889.x
Rebel, H. 1934. Bernstein-Lepidopteren (aus der Sammlung Bachofen-Echt). Palaeobiologica, 6:1-16.
Rebel, H. 1935. Bernstein-Lepidopteren. Entomologische Zeitschrift Iris, 49:162-186.
Regier, J.C., Mitter, C., Davis, D.R., Harrison, T.L., Sohn J.-C., Cummings, M.P., Zwick, A. ja Mitter, K.T. 2014. A molecular phylogeny and revised classification for the oldest ditrysian moth lineages (Lepidoptera: Tineoidea), with implications for ancestral feeding habits of the mega-diverse Ditrysia. Systematic Entomology, 40(2). https://doi.org/10.1111/syen.12110
Scoble, M.J. 1995. Lepidoptera – muoto, toiminta ja monimuotoisuus. The Natural History Museum / Oxford University Press, Oxford.
Skalski, A.W. 1973. Lepidoptera-tutkimuksia fossiilisista hartseista. Osa II. Epiborkhausenites obscurotrimaculatus gen. et sp. nov. (Oecophoridae) ja Baltian meripihkasta löydetty Tineid-perhonen. Acta Palaeontologica Polonica, 18(1):153-160.
Skalski, A.W. 1974. Zwei neue Gattungen und Arten der Familie Tineidae aus dem Baltischen Bernstein. Studien an Lepidopteren aus fossilen Harzen V. Beiträge zur Entomologie, 1/4(24):97-104.
Skalski, A.W. 1977. Lepidoptera-tutkimuksia fossiilisista hartseista. Osa I. Yleisiä huomioita ja kuvauksia uusista sukujen Tineidae ja Oecophoridae sukujen suvuista ja lajeista balttilaisesta meripihkasta. Prace Museum Ziemi, 26:3-24.
Sohn, J.-C., Labandeira, C., Davis, D., and Mitter, C. 2012. An annotated catalog of fossil and subfossil Lepidoptera (Insecta: Holometabola) of the world. Zootaxa, 3286:1-132.
Sohn, J.-C. ja Lamas, G. 2013. Corrections, additions, and nomenclatural notes to the recently published world catalog of fossil and subfossil Lepidoptera. Zootaxa, 3599:395-399.
Sontag, E. 2003. Eläinsulkeumat valikoimattomasta balttilaisesta meripihkasta otetussa näytteessä. Acta Zoologica Cracoviensia, 46 (suppl.-Fossil Insects):431-440.
van Nieukerken, E.J., Kaila, L., Kitching, I.J., Kristensen, N.P., Lees, D.C., Minet, J., Mitter, C., Mutanen, M., Regier, J.C., Simonsen, T.J., Wahlberg, N., Yen, S.-H., Zahiri, R., Adamski, D., Baixeras, J., Bartsch, D., Bengtsson, B.Å., Bengtsson, B.Å., Brown, J.W., Bucheli, S.R., Davis, D.R., Davis, D.R., De Prins, J., De Prins, W., Epstein, M.E., Gentili-Poole, P., Gielis, C., Hätteschwiler, P., Hausmann, A., Holloway, J.D., Kallies, A., Karsholt, O., Kawahara, A.Y., Koster, S.J.C., Kozlov, M.V., Lafontaine, J.D., Lamas, G., Landry, J.-F.., Lee, S., Nuss, M., Park, K.-T., Penz, C., Rota, J., Schintlmeister, A., Schmidt, B.C., Sohn, J.-C., Sohn, J.-C., Solis, M.A., M.A., Tarmann, G.M., Warren, A.D., Weller, S., Yakovlev, R.V., Zolotuhin, V.V., and Zwick, A. 2011. Lepidoptera Linnaeus, 1758. Teoksessa Zhang, Z.-Q. (toim.), Eläinten biologinen monimuotoisuus: An outline of higher-level classification and survey of taxonomic richness. Zootaxa, 3148:212-221.
von Linné, C. 1758. kääntänyt William Turton (1806). Nide 1. A General System of Nature: Through the Three Grand Kingdoms of Animals, Vegetables, and Minerals, Systematically Divided into their Several Classes, Orders, Genera, Species, and Varieties. Lackington, Allen, and Co, London.
Wagner, D.L. ja Rosovsky, J. 1991. Mating systems in primitive Lepidoptera, with emphasis on the reproductive behavior of Korscheltellus gracilis (Hepialidae). Zoological Journal of the Linnean Society, 102:277-303. https://doi.org/10.1111/j.1096-3642.1991.tb00003.x
Weitschat, W. ja Wichard, W. 2002. Atlas Itämeren meripihkan kasveista ja eläimistä. Verlag Friedrich Pfeil, München.
Weitschat, W. 2009. Jäger, Gejagte, Parasiten und blinde Passagiere – Momentaufnahmen aus dem Bernsteinwald. Denisia, 26(86):243-256.
Wichard, W., Gröhn, C. ja Seredszus, F. 2009. Aquatic Insects in Baltic Amber. Verlag Kessel, Remagen-Oberwinter.