- Párzó lepkék (Tineidae, Ditrysia, Lepidoptera) fagyasztott viselkedési zárványként konzerválódtak a balti borostyánban (eocén)
- ABSTRACT
- BEVEZETÉS
- MATERIALIS ÉS MÓDSZEREK
- SYSTEMATIC PALAEONTOLOGY
- DISZKURZUS
- Véletlen együttes előfordulás vagy megőrzött párzás?
- Taphonomia
- Szexuális dimorfizmus
- Habitat
- ÖSSZEGZÉS
- FELHÍVÁSOK
Párzó lepkék (Tineidae, Ditrysia, Lepidoptera) fagyasztott viselkedési zárványként konzerválódtak a balti borostyánban (eocén)
Thilo C. Fischer és Marie K. Hörnig
Cikkszám: 22.1.7
https://doi.org/10.26879/829
Copyright Paleontological Society, February 2019
Autori életrajzok
Szöveges és többnyelvű összefoglalók
PDF változat
Elküldés: 2017. október 22. Acceptance: 23 Januiary 2019
{flike id=2421}
ABSTRACT
A borostyán zárványok vizsgálata egyedülálló betekintést nyújt az évmilliókkal ezelőtt élt állatok biológiájába, mivel kivételes, szinte életszerű körülmények között megőrződtek. Ez nem csak a morfológiai szempontokra igaz, a viselkedés aspektusaira is lehet következtetni ezekből a fosszíliákból.
Itt az eocén balti borostyánból származó Microlepidopterákról számolunk be, valószínűleg a kopulában talált fosszilis Lepidopterák első előfordulásaként. A lepkék a Tineidae családba tartoznak (a Tineidae lepkékhez tartoznak a ruhamolyok), és új fajként és új nemzetségként, Forcepsites michalskii néven írják le őket. A hímek kapaszkodó szerve meglehetősen szokatlan a balti borostyánból származó lepkéknél, egy ventrális és egy dorzális részből áll, mindegyik két párhuzamos, tövisszerű kiemelkedésben végződik, és a jelek szerint nem a vápák alkotják. A hím és a nőstény együttes előfordulása, valószínűleg kopulában, lehetővé tette szexuális dimorfizmusuk közvetlen tanulmányozását, és bizonyíték van arra, hogy a faj élőhelye a balti borostyánerdőben a gyantatermelés helyszíne. Az antennák és a szemmorfológia nemi dimorfizmusából értelmezhető, hogy a nőstény volt a párzásra hívó nem ebben a fajban.
Thilo C. Fischer. Förderverein der Bayerischen Staatssammlung für Paläontologie und Historische Geologie München Richard-Wagner-Straße 10, D-80333 München, Németország.
Marie K. Hörnig. Greifswaldi Egyetem, Állattani Intézet és Múzeum, citológia és evolúcióbiológia, Soldmannstr. 23, D-17489 Greifswald, Németország.
FÜGGŐSZÓK: Ruhamolyok; élőhely a kopulában; Microlepidoptera; új fajok és nemzetségek; szexuális dimorfizmus
Végleges idézet: Fischer, Thilo C. és Hörnig, Marie K. 2019. Párzási lepkék (Tineidae, Ditrysia, Lepidoptera) fagyasztott viselkedéses felvételként megőrződve a balti borostyánban (eocén). Palaeontologia Electronica 22.1.7A 1-11. https://doi.org/10.26879/829
palaeo-electronica.org/content/2019/2421-mating-moths
Copyright: Február 2019 Paleontológiai Társaság.
Ez egy nyílt hozzáférésű cikk, amelyet az Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0) feltételei szerint terjesztünk, ami lehetővé teszi a felhasználók számára az anyag másolását és újraelosztását bármilyen médiumban vagy formátumban, feltéve, hogy azt nem használják fel kereskedelmi célokra, és feltüntetik az eredeti szerzőt és forrást, feltüntetve, ha bármilyen változtatás történt.
creativecommons.org/licenses/by-nc-sa/4.0/
http://zoobank.org/BCF64D89-6879-4805-8021-7CE926CE4B12
BEVEZETÉS
A párzó rovarok ritkán fordulnak elő borostyánban az úgynevezett “fagyott viselkedés” eseteiként (Arillo, 2007 és hivatkozások). Ilyeneket jelentettek az eocén balti borostyánból, például a Diptera, Coleoptera, Hemiptera, Hymenoptera és meglehetősen ritkán a Trichoptera csoportokból (Weitschat és Wichard, 2002; Weitschat, 2009; Boucot és Poinar Jr., 2010; Gröhn, 2016).
A balti borostyánban a Lepidoptera általában ritka, és körülbelül minden 250 zárványból egy fordul elő (összehasonlításképpen: A balti borostyánban a Dipterák a rovarok kb. 64%-át, a Lepidopterák kb. 0,4%-át képviselik; Hoffeins és Hoffeins 2003; Gröhn 2016). Más rovarokhoz hasonlóan a kopulában megőrződött imágók sokkal ritkábbak, és kopulában élő lepkeszúnyogokat még nem írtak le, még más borostyánból vagy nem borostyán rovarlelőhelyekről sem (Sohn et al., 2012; Sohn és Lamas, 2013). Az in copula előfordulása lehetővé teszi mindkét ivaros forma jóhiszeműen egymáshoz, illetve mindkettő együttesen egy külön fajhoz való tartozását, és így egy faj ivaros dimorfizmusának pontos leírását. Alternatívaként a fosszilis rovarok szexuális dimorfizmusát csak statisztikai módszerekkel lehet vizsgálni, amikor egy faj leletei bőségesek, mint például számos Nematocera esetében (pl, Borkent, 1995).
A Tineidae a Ditrysia lepidoptera csoport erősen divergens bazális képviselői (Regier et al., 2014), lárváik a szubsztrátok széles skálájával (detritus, gombák, zuzmók vagy keratin) táplálkoznak (Scoble, 1995), ezért generalisták (85,7%) (Menken et al., 2009). Balti borostyán zárványokként a Tineidae az összes lepidoptera lelet mintegy negyedét képviselik (Skalski, 1977). Itt a Tineidae egy ritka és új faját írjuk le ivaros formáival együtt egy feltételezett kopulában előforduló előfordulásból.
MATERIALIS ÉS MÓDSZEREK
A borostyánmintát Artur Michalski (Wroclaw, Lengyelország) találta és vásárolta meg. Akrillakkal történő kezeléssel konzerválták, és a szerző (TF) gyűjteményéből származik, gyűjtési száma 7131. A tárolás állandó hőmérsékleten, műanyag klipszes zacskókban, fémdobozokban történt, oxigén és fény kizárásával. A példányt a müncheni Bayerische Paläontologische Staatssammlung Münchenben helyezik letétbe SNSB-BSPG 2017 I 113.
A példányt Canon MP-E 65 mm-es objektívvel és Canon MT-24 Twin Flash vakuval felszerelt Canon EOS 70d fényképezőgéppel fényképezték. A tükröződések elkerülése érdekében a vaku és a fényképezőgép objektívje elé polarizációs szűrőket szereltünk és állítottunk be (keresztpolarizált fény). A borostyánfelület domborúságának és homorúságának csökkentése érdekében egy csepp glicerint alkalmaztunk, és fedőpapírral fedtük le. A felvett kép z-halmazait Combine ZP-vel fuzionáltuk, majd Adobe Photoshop CS 3 programmal tovább dolgoztuk fel. A zárványt mikrokomputer-tomográfiával nem lehetett vizsgálni, mert a fémszerű, piritnek tűnő zárványok jelenléte erős leleteket okoz.
SYSTEMATIC PALAEONTOLOGY
Systematics (van Nieukerken et al. szerint, 2011)
Rend LEPIDOPTERA Linnaeus, 1758
Clade DITRYSIA Börner, 1925
Superfamily TINEOIDEA Latreille, 1810
Family TINEIDAE Latreille, 1810
Genus Forcepsites nov. gen.
zoobank.org/DDB6884D-845A-4A87-981E-A2953ADD8964
Etymology. “Forceps”, latinul fogó, a hím fogószervére utalva.
Fajok Forcepsites michalskii nov. spec.
zoobank.org/5494D58E-7773-473C-AFB2-140DDD049226
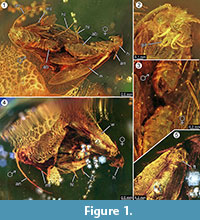 Holotypus. Specimen ex coll. Fischer no. 7131, 1.2. ábra
Holotypus. Specimen ex coll. Fischer no. 7131, 1.2. ábra
Locus typicus. Yantarni borostyánbánya, GUS
Stratum typicum. “Blaue Erde” (felső eocén – alsó oligocén)
Etimológia. A michalskii fajnevet Artur Michalski elismeréseként választottuk, aki ezt a példányt biztosította.
Tárhely. Bayerische Staatssammlung für Paläontologie und Geologie, München, Németország; hozzáférési szám: SNSB-BSPG 2017 I 113.
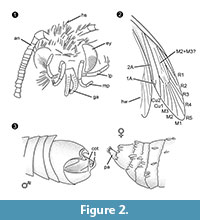
A nemzetség diagnózisa. Monotipikus nemzetség, amelynek meghatározó jellemzője a hímek kapaszkodó szerve (2.3. ábra), amelynek ventrális és dorzális része két párhuzamos, tövisszerű kiemelkedésben végződik.
 A faj diagnózisa. A nőstény és hím imágók (1. ábra) megfelelő minőségi és mennyiségi jellemzőit határoztuk meg, amelyeket az 1. táblázatban foglaltunk össze.
A faj diagnózisa. A nőstény és hím imágók (1. ábra) megfelelő minőségi és mennyiségi jellemzőit határoztuk meg, amelyeket az 1. táblázatban foglaltunk össze.
Differenciáldiagnózis. A karakterek kombinációja kis proboscis különálló galeákkal, hosszú és összehajtott maxilláris szájpadok öt szegmenssel, felfelé mutató és enyhén hajlított labialis szájpadok oldalsó sörtékkel, fej sok felálló szőrszerű pikkellyel erősen a Tineidae-hoz való tartozást sugallja, de ezek egyike sem apomorf, és ilyen apomorfia jelenleg sem ismert a kifejlett Tineidae esetében (Regier et al., 2014). Az R4 és R5 erek nem összenőttek/szárasak, ami a Gelechioidea (Gelechiidae és Oecophoridae) (Scoble, 1995), a balti borostyánban talált Lepidoptera másik nagy csoportjára utalna. A faj nem hasonlít a Tineidae vagy más családok leírt képviselőihez a balti borostyánból (Rebel, 1934, 1935; Kuznezov, 1941; Skalski, 1973; Kozlov, 1987, 1988; Sohn et al., 2012; Sohn és Lamas, 2013), különösen a hímivarszervek tekintetében. Leginkább hasonlónak tűnik a Tineosemopsis decurtatus Skalski 1974 (nőstény ismert), de különbözik a felálló szőrök hiányában a csúcson és a kisebb szemekben. A Tillyardinea eocaenica Kusnezov 1941 (nőstény), amely a csúcson felálló pikkelyekkel és hasonló erezettséggel rendelkezik, a fronok és a szájpadok tekintetében különbözik. A Simulotinea intermedia Skalski 1977 habitusa hasonló, de különbözik egy elágazó véna jelenlétében is, amely a mellső szárny nagy sejtjét osztja (M2+3).
Táfonomia. A Forcepsites michalskii hímje és nősténye egymás mellett fosszilizálódott, és a has hátsó része egymás felé irányul. A két zárvány egy 15 mm átmérőjű és 11 mm magas, nem autoklávozott balti borostyán hengeres darabjában található, amelyet magasságának mintegy felénél két gyantafolyós réteg közötti egyenetlen határvonal tagol. Ennek a határnak a síkjában egy piritréteg látható. Az egyik gyantafolyás tartalmazza a két molylepkét, amelyek hátoldalukat ennek a határnak kiteszik. A női szárny pereméről származó hosszú pikkelyek megmaradtak. A nőstény imágó két lábát elvesztette, és a mellkasa sérült. A másik folyás nem lineáris buboréksorozatot tartalmaz. Mindkét egyed frissen kifejlődöttnek tűnik, amint azt a szárnyakon lévő pikkelyek teljes mellénye és a szárnyak nem foszlós volta alapján meg lehet ítélni. A szárnyakon, a hason és a lábakon nem lehetett illatszerveket azonosítani. A Microlepidoptera egyedeknél csak a balti borostyánra jellemző trichomák maradtak fenn.
DISZKURZUS
Véletlen együttes előfordulás vagy megőrzött párzás?
A mellékelt lepkék ugyanannak a fajnak a hím és nőstény képviselői, és egymás mellett helyezkednek el, a hasi részek egymás felé tájolva. A lepkék nemi szervei mégsem közvetlenül érintkeznek egymással, hanem egymáshoz közel helyezkednek el. A hím és a nőstény dorzális/ventrális tájolása kissé eltér, amit a testtengelyen belüli enyhe forgás okoz (különösen a hímnél). Mindkét lepke hátsó hasi részének orientációja egymás felé irányul.
Már több olyan eset is ismert, amikor a még folyékony gyantába való bezártság helyzete stresszreakciót, például peteérést idézhet elő (Weitschat, 2009). A párzási viselkedés indukálása a gyanta áramlása általánosságban nem hihető, de a bezártsági helyzet azt mutatja, hogy a friss gyantán belüli enyhe mozgások lehetségesek voltak.
Felmerül az a kérdés is, hogy ez a megállapítás jelenthet-e véletlen együttes előfordulást is. Egy ilyen véletlen együttes előfordulásának valószínűsége nagyjából megbecsülhető. A rovarzárványok közül a molyok a balti borostyánban 250 zárványból körülbelül egy gyakorisággal fordulnak elő (Krzeminskaet al., 1991 (0,6%); Hoffeins és Hoffeins, 2003 (0,4%); Sontag, 2003 (0,27%) és saját megfigyelés). Egy egyedi borostyánkő esetében, amelyben egy moly van, a másik moly véletlenszerű együttes előfordulásának esélye egyszerűen 1:250 (a molyok nem fordulnak elő rajokban). Ugyanannak a taxonnak, vagy egy olyan bőséges taxonnak az együttes előfordulása, amely tévedésbe eshet, a legjobb esetben is 1:10-re becsülhető (Skalski, 1977), és még körülbelül 1:2 a különböző nemek megléte esetén. Figyelembe véve a szoros elrendeződés további esélyeit, mind a testtengelyek összehangolását, mind a megfelelő testorientációt, a párzási eseményként tévesen értelmezhető véletlenszerű együttes előfordulás valószínűsége jóval kisebb, mint a vizsgált szerző gyűjteményében a 920 lepidoptera-besorolások közül egynek a megfigyelt gyakorisága.
Végeredményben ennek az eredménynek a legvalószínűbb értelmezése az, hogy a lepkék kopulában voltak, és a genitáliák érintkeztek, de az egyedek a gyantaáramlás miatt kissé eltávolodtak egymástól (lásd az alábbi tárgyalást).
Taphonomia
A gyantafolyások gondos elemzésének jelentőségét a borostyánbefogások taphonomiája szempontjából Coty és munkatársai (2014) bizonyították a mexikói Chiapas borostyánban megőrződött hangya – termesz interakció vizsgálatával.
A kopulában történő esemény előfordulását erősen alátámasztja a nőstény szárny pereméről származó hosszú pikkelyek megtalálása, amelyek bizonyos távolságra a hím irányába sodródtak. Ez egy erős lamináris gyantaáramlásra utalhatott ezen a helyen, amely a közös tengelyük mentén mozgott, és a most megfigyelt rövid távolságra távolíthatta el őket egymástól. A lepkék párzása során a hím elveszítheti a kapcsolatot a párzási hellyel, és előfordulhat, hogy csak a kapaszkodó szerve rögzíti a nőstényhez, és csak a nőstény érintkezik a felszínnel. Ez a viselkedés és a hirtelen gyantaáramlás okozta vonszolás lehet az oka annak, hogy a nőstény elvesztette mindkét hátsó lábát. A lepkék hátoldalán, amely ennek az első lamináris gyantaáramlásnak a domború felületén volt kitéve, a nőstény szárnya és az alatta lévő mellkas részben sérült (1.5. ábra). Feltételezhetően ezek a roncsolódott területek más rovarok, valószínűleg hangyák által okozott táplálkozási nyomokat jelentenek, amint az a borostyánmintákon gyakran megfigyelhető. Ezért az első gyantafolyás mint beágyazási esemény után a háti oldalak egy ideig szabadon voltak, elég hosszú ideig ahhoz, hogy a ragadozók ezen a szilárd felületen járhassanak. A második gyantaáramlás, amely később borította be ezt az oldalt, gyors és turbulens gyantaáramlás lehetett, amint azt a benne található szabálytalan görbékbe rendezett kis buborékok sorozata jelzi. Ez tipikus taphonomiai folyamat lenne egy úgynevezett “Schlaube” balti borostyán esetében (Gröhn, 2016).
Szexuális dimorfizmus
A szexuális dimorfizmust nehéz megfigyelni a fosszilis fajoknál, amelyekből általában csak néhány példány ismert. A nem által nem befolyásolt karaktereknek diagnosztikusnak kell lenniük ahhoz, hogy a nemek további megkülönböztetésének és a szexuális dimorfizmus tisztázásának alapjaként biztos fajmeghatározást tegyenek lehetővé. Tekintettel az erősen nemi dimorfikus élő fajokra, pl. a brachypterus nőstényekre, mint sok Psychidae esetében (Scoble, 1995), a két nem egy fajhoz való tartozására való következtetés aligha lenne lehetséges. Ezért csak a gyenge szexuális dimorfizmus esetei ismerhetők fel megbízhatóan a fosszilis fajokban statisztikai eszközökkel és anélkül, hogy a kopulában megtalálnánk őket. Ez aláhúzza a fosszilis fajok azonosított párzási viselkedésének fontosságát az erős szexuális dimorfizmus ilyen eseteinek leírásához. A fosszilis párosodás ilyen megőrzött leletei jóváhagyhatják a korábban ismert szexuálisan dimorf formák egy fajhoz tartozásának korrelációját is, még a nagyfokú statisztikai alátámasztás lehetősége nélkül is. Van némi esély arra, hogy egy megőrzött párosodási esemény fajok közötti vagy nemek közötti párosodást képviselhet, mivel ezek ismertek lepkékről és pillangókról (Ganyard és Brady, 1972; Deering és Scriber, 2002). Ezek a jelenségek azonban sokkal ritkábbak, mint a fajon belüli párosodások, és aligha valószínű, hogy ritka fosszilis felvételként megtalálhatók.
A Forcepsites michalskii-nál megfigyelt nem allometrikus nemi dimorfizmus a hímeknél a proboszkuszban levő elkülönített galeák. Ez egy nem funkcionális állapotra utalhat, és e faj hím imágói nem táplálkoznak, ahogyan azt számos, csökevényes vagy hiányzó proboszkizmussal rendelkező (hím) Psychidae-nél láthatjuk (Scoble, 1995). Az itt talált allometrikus nemi dimorfizmusok a következők: (1) A nőstény nagyobb, mint a hím, mind testméretben, mind szárnyhosszban. A vonatkozó viszonyok azonban közel azonosak. (2) A nősténynél a has viszonylag és abszolút szélesebb a hímhez képest. (3) A hím csápjai sokkal nagyobbak és vastagabbak, mint a nőstény imágónál. Számos ma élő fajnál az antennák nemileg dimorfak (Scoble, 1995), a jól fejlett hím antennák a nőstény feromonok kemoszenzorálására szolgálnak. (4) A hímek maxilláris tapogatói kétszer olyan vastagok, mint a nőstényeké, és bordás szerkezetűek. (5) A hímek szemei szintén nagyobbak, de az ommatídiumok száma körülbelül ugyanannyi. A nagyobb szemek csak több fényt fogadnak be, és érzékenyebbek lehetnek, a felbontás azonban nem változik.
A hívó nőstény lepkék hosszú távú felismerése a hím kemoreceptorain keresztül történik, míg a rövid távú felismerés ehelyett a vizuális azonosításra támaszkodik (Groot, 2014). A Forcepsites michalskii esetében is úgy tűnik, hogy a nőstény volt a hívó nem -mire utal a hím nagyobb antennakészüléke és szeme is.
Habitat
A fent bemutatott értekezés alapján egyértelmű, hogy a kopulában fogott egyedek csak ott fordulhatnak elő, ahol ez a faj élt és párosodott. Az egyes egyedeket elsodorhatja a szél, vagy felkereshetik az élőhelyen kívüli helyeket, mint a balti borostyánban ritkán előforduló vízi rovarok (Wichardet al., 2009); a párzó egyedek esetében viszont ez aligha történhetett meg anélkül, hogy szétváltak volna. Úgy tűnik, a szakirodalomban nincs bizonyíték a lepkék repülés közbeni párosodására. Ezzel szemben a legtöbb Lepidoptera és a túlnyomórészt éjszaka élő mikrolepidopterák (lepkék) esetében a nőstények feromonok segítségével “hívogatják” a hímeket, így határozzák meg a párzási helyeket (Groot, 2014). Előfordulhat, hogy a párzás előtt repülnek, de a legtöbb esetben úgy tűnik, hogy ülő helyzetben vannak, és a feromonok terjesztése érdekében legyeznek. Egyes taxonoknál azonban a hívó vagy kereső ivarok fordítottak is lehetnek, de a Tineidae esetében a jelentések szerint a nőstény a hívó ivar (Wagner és Rosovsky, 1991; Scoble, 1995). Mindenesetre a gyantafolyás helye, azaz a borostyántermelő fákat tartalmazó biotóp valószínűleg az itt leírt ruháslepke élőhelye is volt.
ÖSSZEGZÉS
Eocén balti borostyánból származó új faj és nemzetség Forcepsites michalskii néven egy figyelemre méltó hímivarszervi apparátussal rendelkező tineoid lepkét írunk le. A nőstény és a hím együtt, szoros helyzetben és egymás felé tájolt hasüreggel maradt fenn, ami arra utal, hogy ezek a lepkék a párzás során a friss gyantában rekedtek. Egy erős lamináris gyantaáramlás, amelyre a nőstény testének több elsodródott része is utal, távolíthatta őket egymástól a most megfigyelt rövid távolságra. Ez a példány tehát a kopulában befogott fosszilis lepkék első példája lehet.
A Forcepsites michalskii feltételezhetően erdőlakó faj volt, mivel kopulában befogott egyedek csak arról a helyről dokumentálhatók, ahol ez a faj élt és párosodott.
A F. michalskii nőstényei és hímjei nemi dimorfizmust mutatnak a testméret, a szárnyhossz, a has arányai, az összetett szemek mérete és a maxilláris tapogatók, valamint az antennák morfológiája tekintetében. A hím egyértelműen nagyobb és vastagabb antennái, valamint nagyobb szemei alapján azt feltételezzük, hogy a nőstény lehet a hívó nem ebben a fajban, ahogyan azt a Tineidae fajokról is jelentették.
FELHÍVÁSOK
A. Michalskinak (Wroclaw, Lengyelország) köszönjük, hogy ezt a kivételes példányt rendelkezésünkre bocsátotta és felhívta rá a figyelmünket. M. Schwarz az Oberösterreichisches Landesmuseum (Linz, Ausztria) Lepidoptera gyűjteményének átvizsgálását szívesen támogatta. MKH a Studienstiftung des deutschen Volkes ösztöndíjával kedveskedett, és köszönetet mond S. Harzschnak (Greifswald, Németország) a támogatásért. Köszönjük M. Heikkilä és C. Haug (München, Németország) értékes tudományos észrevételeit a kézirathoz, valamint J. Krieger (Greifswald, Németország) megjegyzéseit és C. MacGraphery (Freising, Németország) és Y. Goldman (Collinsville, CT, USA) anyanyelvi támogatását az angol szöveg ellenőrzésében. Továbbá köszönetet mondunk két névtelen bírálónak a hasznos megjegyzéseikért.
Arillo, A. 2007. Paleoetológia: megkövült viselkedésformák borostyánban. Geologica Acta, 5(2):159-166. https://doi.org/10.1344/105.000000301
Börner, C. 1925. Lepidoptera, Schmetterlinge, p. 358-387. In Brohmer, P. (szerk.). Fauna von Deutschland. Ein Bestimmungsbuch unserer heimischen Tierwelt. Quelle und Meyer, Leipzig.
Borkent, A. 1995. Csípőszúnyogok Észak-Amerika kréta borostyánjában (Diptera: Ceratopogonidae). Backhuys Publishers, Leiden, Hollandia.
Boucot, A.J. és Poinar Jr, G.O. 2010. Fossil Behavior Compendium. CRC Press, Boca Raton, p. 147-151.
Coty, D., Aria, C., Garrouste, R., Wils, P., Legendre, F. és Nel, A. 2014. Az első hangya-termita szininklúzió borostyánban, CT-szkenneres taphonomiai elemzéssel. PLOS one, 9(8):e104410. https://doi.org/10.1371/journal.pone.0104410
Deering, M.D. és Scriber, J.M. 2002. Terepi biotesztek heterospecifikus párzási preferencia aszimmetriát mutatnak a hibridizáló észak-amerikai Papilio lepkefajok (Lepidoptera: Papilionidae) között. Journal of Ethology 20:25-33. https://doi.org/10.1007/s10164-002-0050-2
Fletcher, T.B. 1940. Új nemzetségnevek a mikrolepidopterákhoz. Entomologist’s Record, 52:17-19.
Ganyard, M.C. és Brady, Jr.U.E. 1972. Interspecifikus vonzódás Lepidopteráknál a terepen. Annals of the Entomological Society of America, 65 (6): 1279-1282. https://doi.org/10.1093/aesa/65.6.1279
Gröhn, C. 2016. A balti borostyánban található zárványok. Wachholtz Verlag – Murmann Publishers, Kiel, Hamburg.
Groot, A.T. 2014. A lepkék szexuális tevékenységének cirkadián ritmusa: Áttekintés. Frontiers in Ecology and Evolution 2:43. https://doi.org/10.3389/fevo.2014.00043
Hoffeins, C. és Hoffeins, H.W. 2003. Vizsgálatok a nem válogatott gyűjtésekből származó balti és bitterfeldi borostyán (harmadidőszak, eocén) zárványainak gyakoriságáról, különös tekintettel a Diptera rendre. Studia Dipterologica, 10(2):381-392.
Kozlov, M.V. 1987. Új lepkeszerű lepkék a balti borostyánból. Paleontological Journal, 4:59-67.
Kozlov, M.V. 1988. A Lepidopterák paleontológiája és a Papilionida rend filogenetikájának problémái, p. 16-69. In Ponomarenko A.G. (szerk.), The Mesozoic-Cenozoic Crisis in the Evolution of Insects. Academy of Sciences, Moscow.
Krzeminska, E., Krzeminski, W., Haenni, J.-P., and Dufour, C. 1991. Les Fantomes De L’Ambre. Muséed’histoire naturelle de Neuchâtel, Neuchâtel, Svájc.
Kusnezov, N. 1941. A borostyánlepkék felülvizsgálata. Paleontológiai Intézet, Szovjetunió Tudományos Akadémiája, Moszkva & Leningrád.
Latreille, P.A. 1810. Általános megfontolások a rákfélék, pókfélék és rovarok osztályait alkotó állatok természetes rendjéről: családokba rendezett nemzetségek módszertani táblázatával. Paris, Schoell.
Menken, S.B.J., Boomsma, J.J. és van Nieukerken, E.J. 2009. A gazdanövénytársulások nagyléptékű evolúciós mintázatai a Lepidopteráknál. Evolution, 64(4):1098-1119. https://doi.org/10.1111/j.1558-5646.2009.00889.x
Rebel, H. 1934. Bernstein-Lepidopteren (aus der Sammlung Bachofen-Echt). Palaeobiologica, 6:1-16.
Rebel, H. 1935. Bernstein-Lepidopteren. Entomologische Zeitschrift Iris, 49:162-186.
Regier, J.C., Mitter, C., Davis, D.R., Harrison, T.L., Sohn J.-C., Cummings, M.P., Zwick, A., and Mitter, K.T. 2014. A legrégebbi ditriai lepkefajok (Lepidoptera: Tineoidea) molekuláris filogeniája és felülvizsgált osztályozása, a mega-diverz Ditrysia ősi táplálkozási szokásaira vonatkozó következtetésekkel. Systematic Entomology, 40(2). https://doi.org/10.1111/syen.12110
Scoble, M.J. 1995. A Lepidoptera – forma, funkció és sokféleség. The Natural History Museum / Oxford University Press, Oxford.
Skalski, A.W. 1973. Tanulmányok a Lepidopterákról fosszilis gyantákból. Part II. Epiborkhausenites obscurotrimaculatus gen. et sp. nov. (Oecophoridae) és egy balti borostyánban felfedezett Tineid-moly. Acta Palaeontologica Polonica, 18(1):153-160.
Skalski, A.W. 1974. Zwei neue Gattungen und Arten der Familie Tineidae aus dem Baltischen Bernstein. Studien an Lepidopteren aus fossilen Harzen V. Beiträge zur Entomologie, 1/4(24):97-104.
Skalski, A.W. 1977. Tanulmányok a Lepidopterákról fosszilis gyantákból. I. rész: Általános megjegyzések és a Tineidae és Oecophoridae családok új nemzetségének és fajainak leírása balti borostyánból. Prace Museum Ziemi, 26:3-24.
Sohn, J.-C., Labandeira, C., Davis, D., and Mitter, C. 2012. An annotated catalog of fossil and subfossil Lepidoptera (Insecta: Holometabola) of the world. Zootaxa, 3286:1-132.
Sohn, J.-C. and Lamas, G. 2013. Javítások, kiegészítések és nevezéktani megjegyzések a fosszilis és szubfosszilis lepkeszúnyogok nemrég megjelent világkatalógusához. Zootaxa, 3599:395-399.
Sontag, E. 2003. Állati zárványok egy nem szelektált balti borostyán mintában. Acta Zoologica Cracoviensia, 46 (suppl.-Fossil Insects):431-440.
van Nieukerken, E.J., Kaila, L., Kitching, I.J., Kristensen, N.P., Lees, D.C., Minet, J., Mitter, C., Mutanen, M., Regier, J.C., Simonsen, T.J., Wahlberg, N., Yen, S.-H., Zahiri, R., Adamski, D., Baixeras, J., Bartsch, D., Bengtsson, B.Å., Brown, J.W., Bucheli, S.R., Davis, D.R., De Prins, J., De Prins, W., Epstein, M.E., Gentili-Poole, P., Gielis, C., Hätteschwiler, P., Hausmann, A., Holloway, J.D., Kallies, A., Karsholt, O., Kawahara, A.Y., Koster, S.J.C., Kozlov, M.V., Lafontaine, J.D., Lamas, G., Landry, J.-F.,.., Lee, S., Nuss, M., Park, K.-T., Penz, C., Rota, J., Schintlmeister, A., Schmidt, B.C., Sohn, J.-C., Solis, M.A., Tarmann, G.M., Warren, A.D., Weller, S., Yakovlev, R.V., Zolotuhin, V.V., and Zwick, A. 2011. Lepidoptera rend Linnaeus, 1758. In Zhang, Z.-Q. (szerk.), Animal biodiversity: A magasabb szintű osztályozás vázlata és a taxonómiai gazdagság felmérése. Zootaxa, 3148:212-221.
von Linné, C. 1758. fordította William Turton (1806). 1. kötet. A természet általános rendszere: Az állatok, növények és ásványok három nagy birodalmán keresztül, szisztematikusan felosztva különböző osztályaikra, rendjeikre, nemeikre, fajaikra és fajtáikra. Lackington, Allen, and Co, London.
Wagner, D.L. és Rosovsky, J. 1991. Párzási rendszerek a primitív lepkéknél, különös tekintettel a Korscheltellus gracilis (Hepialidae) szaporodási viselkedésére. Zoological Journal of the Linnean Society, 102:277-303. https://doi.org/10.1111/j.1096-3642.1991.tb00003.x
Weitschat, W. és Wichard, W. 2002. Atlas of Plants and Animals in Baltic amber. Verlag Friedrich Pfeil, München.
Weitschat, W. 2009. Jäger, Gejagte, Parasiten und blinde Passagiere – Momentaufnahmen aus dem Bernsteinwald. Denisia, 26(86):243-256.
Wichard, W., Gröhn, C., and Seredszus, F. 2009. Vízi rovarok a balti borostyánban. Verlag Kessel, Remagen-Oberwinter.