Parningsfjärilar (Tineidae, Ditrysia, Lepidoptera) bevarade som frysta beteenden i baltisk bärnsten (Eocen)
Thilo C. Fischer och Marie K. Hörnig
Artikelnummer: 22.1.7
https://doi.org/10.26879/829
Copyright Paleontological Society, February 2019
Författarbiografier
Plåspråkiga och flerspråkiga sammanfattningar
PDF-version
Inlämning: 22 oktobere 2017. Acceptans: 23 januari 2019
{flike id=2421}
ABSTRACT
Undersökningen av bärnstensinklusioner ger en unik inblick i biologin hos djur som levde för miljontals år sedan, på grund av deras exceptionella bevarande i nästan livslika förhållanden. Detta gäller inte bara morfologiska aspekter, aspekter av beteende kan också härledas från dessa fossil.
Här rapporteras Microlepidoptera från eocen baltisk bärnsten som sannolikt är den första förekomsten av fossila Lepidoptera som hittats i copula. Mottarna tillhör Tineidae (tineid-motar inkluderar klädmottar) och beskrivs som en ny art och ett nytt släkte, Forcepsites michalskii. Hanens klämorgan är ganska ovanligt för nattfjärilar från baltisk bärnsten, med en ventral och en dorsal del, som var och en slutar i två parallella, taggliknande utbuktningar, och som verkar inte bildas av valvorna. Samförekomsten av hane och hona, troligen i copula, gjorde det möjligt att studera deras sexuella dimorfism direkt, och det finns belägg för att artens livsmiljö är platsen för hartstillverkning i den baltiska bärnstensskogen. Utifrån den sexuella dimorfismen hos antennerna och ögonmorfologin tolkas det som att honan var det kallande könet för parning hos denna art.
Thilo C. Fischer. Förderverein der Bayerischen Staatssammlung für Paläontologie und Historische Geologie München Richard-Wagner-Straße 10, D-80333 München, Tyskland.
Marie K. Hörnig. Universitetet i Greifswald, Zoologiska institutet och museet, cytologi och evolutionsbiologi, Soldmannstr. 23, D-17489 Greifswald, Tyskland.
Nyckelord: Klädmottar; habitat i copula; Microlepidoptera; nya arter och släkten; sexuell dimorfism
Slutligt citat: Fischer, Thilo C, and Hörnig, Marie K. 2019. Parningsfjärilar (Tineidae, Ditrysia, Lepidoptera) bevarade som frysta beteendeinklusioner i baltisk bärnsten (Eocen). Palaeontologia Electronica 22.1.7A 1-11. https://doi.org/10.26879/829
palaeo-electronica.org/content/2019/2421-mating-moths
Copyright: February 2019 Paleontological Society.
Detta är en open access-artikel som distribueras enligt villkoren för Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), vilket tillåter användare att kopiera och distribuera materialet i vilket medium eller format som helst, förutsatt att det inte används för kommersiella ändamål och att originalförfattaren och källan krediteras, med angivelser om eventuella ändringar har gjorts.
creativecommons.org/licenses/by-nc-sa/4.0/
http://zoobank.org/BCF64D89-6879-4805-8021-7CE926CE4B12
INLEDNING
Parande insekter hittas sällan i bärnsten som fall av det s.k. ”frusna beteendet” (Arillo, 2007, och referenser däri). Sådana har rapporterats från eocen baltisk bärnsten, till exempel från grupperna Diptera, Coleoptera, Hemiptera, Hymenoptera och ganska sällan Trichoptera (Weitschat och Wichard, 2002; Weitschat, 2009; Boucot och Poinar Jr, 2010; Gröhn, 2016).
I baltisk bärnsten är Lepidoptera generellt sett sällsynta och förekommer i ungefär en av 250 inklusioner (för jämförelse: Dipteraner utgör cirka 64 % av alla insekter i baltiskt bärnsten, Lepidoptera utgör cirka 0,4 %; Hoffeins och Hoffeins 2003; Gröhn 2016). Liksom hos andra insekter är imagos bevarade i copula mycket ovanligare och Lepidoptera i copula har ännu inte beskrivits, inte ens i andra bärnstensarter eller från insektslokaler som inte är bärnstensarter (Sohn et al., 2012; Sohn och Lamas, 2013). En förekomst i copula gör det möjligt att bona fide hänföra båda könsformerna till varandra och båda tillsammans till en distinkt art, och därmed en exakt beskrivning av den sexuella dimorfismen hos en art. Alternativt kan sexuell dimorfism från fossila insekter endast studeras med statistiska metoder när fynd av en art är rikligt förekommande, vilket är fallet med många Nematocera (t.ex, Borkent, 1995).
Tineidae är starkt divergerande basala representanter för lepidopteragruppen Ditrysia (Regier et al., 2014), deras larver livnär sig på ett brett spektrum av substrat (detritus, svampar, lavar eller keratin) (Scoble, 1995), och är därmed generalister (85,7 %) (Menken et al., 2009). Som inklusioner i baltiskt bärnsten utgör Tineidae ungefär en fjärdedel av alla lepidopterafynd (Skalski, 1977). Här beskrivs en sällsynt och ny art av Tineidae med dess sexuella former från en förmodad in copula-förekomst.
MATERIAL OCH METODER
Bärnstensprovet hittades av och köptes av Artur Michalski (Wroclaw, Polen). Det konserverades genom behandling med ett akryllack och kommer från författarens (TF) samling med samlingsnummer 7131. Förvaringen skedde vid konstant temperatur i plastpåsar i metalllådor, utan syre och ljus. Den kommer att deponeras i Bayerische Paläontologische Staatssammlung München med numret SNSB-BSPG 2017 I 113.
Exemplaret fotograferades med en Canon EOS 70d utrustad med ett Canon MP-E 65 mm objektiv och en Canon Twin Flash MT-24. För att undvika reflektioner monterades och justerades polarisationsfilter framför blixtarna och kamerans objektiv (tvärpolariserat ljus). För att minska bärnstensytans konvexitet och konkavitet applicerades en droppe glycerin och täcktes med ett täckglas. De inspelade z-staplarna smältes samman med Combine ZP och bearbetades ytterligare med Adobe Photoshop CS 3. Inklusionen kunde inte studeras med mikrodatortomografi på grund av förekomsten av metalliska, till synes pyritiska inneslutningar som orsakar starka artefakter.
SYSTEMATIC PALAEONTOLOGY
Systematik (enligt van Nieukerken et al, 2011)
Ordningsföljd LEPIDOPTERA Linnaeus, 1758
Clade DITRYSIA Börner, 1925
Överfamiljen TINEOIDEA Latreille, 1810
Familjen TINEIDAE Latreille, 1810
Genus Forcepsites nov. gen.
zoobank.org/DDB6884D-845A-4A87-981E-A2953ADD8964
Etymology. ”Forceps”, latin för tång, vilket syftar på hanens klämorgan.
Species Forcepsites michalskii nov. spec.
zoobank.org/5494D58E-7773-473C-AFB2-140DDD049226
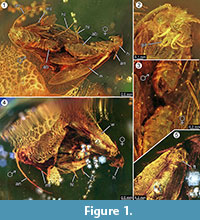 Holotypus. Exemplar ex coll. Fischer nr. 7131, figur 1.2
Holotypus. Exemplar ex coll. Fischer nr. 7131, figur 1.2
Locus typicus. Bärnstensgruva i Yantarni, GUS
Stratum typicum. ”Blaue Erde” (övre Eocen – nedre Oligocen)
Etymologi. Artnamnet michalskii har valts för att uppmärksamma Artur Michalski som tillhandahöll detta exemplar.
Repositorium. Bayerische Staatssammlung für Paläontologie und Geologie, München, Tyskland; accession nummer SNSB-BSPG 2017 I 113.
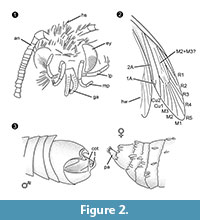
Diagnos av släktet. Monotypiskt släkte med hanens klämorgan (figur 2.3) med en ventral och en dorsal del, som vardera slutar i två parallella, taggliknande utskjutande delar, som definierande kännetecken.
 Diagnos av arten. De respektive kvalitativa och kvantitativa karaktärerna hos de kvinnliga och manliga imagos (figur 1) bestämdes och sammanfattas i tabell 1.
Diagnos av arten. De respektive kvalitativa och kvantitativa karaktärerna hos de kvinnliga och manliga imagos (figur 1) bestämdes och sammanfattas i tabell 1.
Differentialdiagnos. Kombinationen av karaktärerna liten proboscis med separerade galeae, långa och vikta maxillärpalper med fem segment, uppåtriktade och lätt böjda labialpalper med laterala borst, huvudet med många upprättstående hårliknande fjäll tyder starkt på en tillhörighet till Tineidae, men ingen av dessa är apomorfa, inte heller är en sådan apomorfi för närvarande känd för vuxna Tineidae (Regier et al., 2014). Nerverna R4 och R5 är inte sammansmälta/stickade, vilket skulle tyda på Gelechioidea (Gelechiidae och Oecophoridae) (Scoble, 1995), den andra stora gruppen av Lepidoptera som hittats i baltisk bärnsten. Arten skiljer sig från alla beskrivna representanter för Tineidae eller andra familjer från baltisk bärnsten (Rebel, 1934, 1935; Kuznezov, 1941; Skalski, 1973; Kozlov, 1987, 1988; Sohn et al., 2012; Sohn och Lamas, 2013), särskilt när det gäller den manliga genitalapparaten. Mest lik verkar Tineosemopsis decurtatus Skalski 1974 (honan känd) men skiljer sig i avsaknaden av upprättstående hår vid vertex och dess mindre ögon. Tillyardinea eocaenica Kusnezov 1941 (hona), som har upprättstående fjäll vid vertex och en liknande nervering, skiljer sig i fransarna och labialpalperna. Simulotinea intermedia Skalski 1977 har ett likadant habitus men skiljer sig också genom förekomsten av en förgrenad ven, som delar den stora cellen i framvingen (M2+3).
Taphonomi. Hanen och honan av Forcepsites michalskii är fossilerade nära varandra och med bakre delen av buken riktad mot varandra. De två inneslutningarna finns i ett cylindriskt stycke icke-autoklaverad baltisk bärnsten med en diameter på 15 mm och en höjd på 11 mm som är delad på ungefär halva höjden av en ojämn gräns mellan två lager av hartsflöde. Denna kant visar ett lager av pyrit som en del av sitt plan. Ett hartsflöde innehåller de två malarna som exponerar sina respektive ryggsidor mot denna gräns. Långa fjäll som härrör från kanten av honvingarna är bevarade. Honans imago har förlorat två ben och dess thorax är skadat. Det andra flödet innehåller icke-linjära serier av bubblor. Båda individerna verkar vara nyuppkomna, vilket kan bedömas utifrån den kompletta västen av fjäll på vingarna och att vingarna inte är trasiga. Inga doftorgan kunde identifieras på vingar, buk eller ben. De enda syninklusioner som bevarats hos Microlepidoptera är trikomer, typiska för baltiskt bärnsten.
DISKUSSION
Random Co-occurrence or a Preserved Mating?
De bifogade nattfjärilarna är manliga och kvinnliga representanter för samma art och är arrangerade nära varandra med bukarna orienterade mot varandra. Men malarnas genitalier är inte i direkt kontakt, utan i ett nära läge. Den dorsala/ventrala orienteringen hos hane och hona skiljer sig något åt, vilket orsakas av en liten rotation inom kroppsaxeln (särskilt hos hanen). Båda malarnas bakre del av buken är orienterad mot varandra.
Det finns flera kända fall där situationen med inneslutning i den fortfarande flytande hartsen kan framkalla en stressreaktion, t.ex. äggläggning (Weitschat, 2009). Att inducera parningsbeteende genom hartsflödet är generellt sett inte troligt, men situationen med inneslutning visar att små rörelser i den färska hartsen var möjliga.
Det finns också en fråga om detta fynd också skulle kunna representera en slumpmässig samvaro. Sannolikheten för en sådan slumpmässig samförekomst kan grovt uppskattas. Bland insektsinklusioner förekommer malar med en frekvens på ungefär en av 250 inklusioner i baltisk bärnsten (Krzeminskaet al., 1991 (0,6 %); Hoffeins och Hoffeins, 2003 (0,4 %); Sontag, 2003 (0,27 %), och egen observation). Med tanke på en enskild bärnsten där en mal ingår är chansen för slumpmässig förekomst av en annan mal helt enkelt 1:250 (malar förekommer inte i svärmar). Samförekomsten av samma taxon, eller ett rikligt taxon som kan förväxlas, uppskattas i bästa fall till 1:10 (Skalski, 1977), och ytterligare cirka 1:2 för att ha olika kön. Med tanke på de ytterligare chanserna för ett nära arrangemang, både anpassning av kroppsaxlarna och en lämplig kroppsorientering, är sannolikheten för en slumpmässig samförekomst som skulle kunna misstolkas som en parningshändelse mycket lägre än den observerade frekvensen som är en av 920 inklusioner av lepidoptera i den studerade författarens samling.
Den mest sannolika tolkningen av detta fynd är sammanfattningsvis att fjärilarna var i copula och att genitalierna hade varit i kontakt men att individerna var något distraherade från varandra på grund av hartsflödet (se diskussionen nedan).
Taphonomi
Relevansen av en noggrann analys av hartsflöden för taphonomi av bärnstensinklusioner har påvisats av Coty et al. (2014) med sin studie av en interaktion mellan en myra och en termit bevarad i mexikansk Chiapas-bärnsten.
Förekomsten av en in copula-händelse stöds starkt av fyndet av långa skalor som härstammar från kanten av honvingen, som drev en bit i riktning mot hanen. Detta skulle kunna tyda på ett starkt laminärt hartsflöde på denna plats som rörde sig längs deras gemensamma axel och skulle kunna ha dragit isär dem på det korta avstånd som nu observerats. Under parningen av nattfjärilar kan hanen förlora kontakten med parningsplatsen och kan endast vara fäst vid honan med hjälp av klämorganet, och det är endast honan som har kontakt med ytan. Ett sådant beteende och drag från plötsligt hartsflöde kan vara orsaken till att honan har förlorat båda sina bakben. På ryggsidan av fjärilarna, som hade exponerats vid den konvexa ytan av detta första laminära hartsflöde, är honans vinge och dess underliggande bröstkorg delvis skadad (figur 1.5). Man kan anta att dessa förstörda områden representerar födosöksspår orsakade av andra insekter, troligen myror som ofta observeras i bärnstensprover. Efter det första hartsflödet som inbäddningshändelse kommer alltså ryggsidorna att ha varit exponerade under en viss tid, tillräckligt länge för att rovdjur ska kunna gå på denna fasta yta. Det andra hartsflödet, som senare täckte denna sida, kan ha varit ett snabbt och turbulent hartsflöde, vilket indikeras av en serie små bubblor uppställda i oregelbundna kurvor som finns i det. Detta skulle vara en typisk tafonomisk process för en så kallad ”Schlaube” av baltisk bärnsten (Gröhn, 2016).
Könsdimorfism
Könsdimorfism är svår att observera hos fossila arter, från vilka endast ett fåtal exemplar vanligtvis är kända. Karaktärer som inte påverkas av könet måste vara diagnostiska för att möjliggöra en säker artbestämning som grund för ytterligare differentiering mellan könen och belysning av sexuell dimorfism. Med tanke på starkt könsdimorfa existerande arter, t.ex. med brachypterösa honor som hos många Psychidae (Scoble, 1995), skulle det knappast vara möjligt att dra slutsatsen att båda könen tillhör en och samma art. Därför kan endast fall av svag sexuell dimorfism erkännas på ett tillförlitligt sätt i fossila arter med statistiska medel och utan att man hittar dem i copula. Detta understryker vikten av identifierat parningsbeteende hos fossila arter för att beskriva sådana fall av stark sexuell dimorfism. Sådana bevarade fynd av fossil parning kan också godkänna korrelation av tidigare kända sexuellt dimorfiska former att tillhöra en art, även utan möjlighet till mycket statistiskt stöd. Det finns en viss chans att en bevarad parningshändelse kan representera en interspecifik eller intergenerisk parning eftersom dessa är kända från nattfjärilar och fjärilar (Ganyard och Brady, 1972; Deering och Scriber, 2002). Dessa fenomen är dock mycket mindre vanligt förekommande än intraspecifik parning och det är knappast troligt att de hittas som en sällsynt fossil inklusion.
En icke-allometrisk sexuell dimorfism som observerats hos Forcepsites michalskii är de separerade galeae i proboscin hos hanen. Detta kan tyda på ett icke-funktionellt tillstånd, där manliga imagos av denna art inte äter, vilket ses hos många existerande (manliga) Psychidae med rudimentär eller frånvarande proboscis (Scoble, 1995). Allometriska könsdimorfismer som hittats här är: (1) Honan är större än hanen, både i kroppsstorlek och vinglängd. De respektive relationerna är dock nästan lika. (2) Hos honan är buken relativt och absolut bredare i jämförelse med hanen. (3) Hannens antenner är mycket större och tjockare än hos honans imago. Hos flera existerande arter är antennerna sexuellt dimorfa (Scoble, 1995), de välutvecklade hanarnas antenner tjänar till kemosensation av honans feromoner. (4) Hanens maxillära palper är dubbelt så tjocka som hos honans och har en ribbad struktur. (5) Hannens ögon är också större men antalet ommatidia är ungefär detsamma. Större ögon tar bara emot mer ljus och kan vara känsligare; upplösningen skulle dock vara densamma.
Långdistansigenkänning av ropande honfjärilar sker via hanens kemoreceptorer medan kortdistansigenkänning istället förlitar sig på visuell identifiering (Groot, 2014). Hos Forcepsites michalskii verkar det också som om honan var det ropande könet – vilket antyds av hanens större antennapparat och ögon.
Habitat
Baserat på diskussionen som presenterats ovan är det tydligt att individer som fångas i copula endast kan förekomma på den plats där denna art levde och parade sig. Enskilda individer kan drivas med vinden eller besöka platser som inte är livsmiljöer, som vattenlevande insekter som sällan hittas i baltisk bärnsten (Wichardet al., 2009); för parningsindivider i stället skulle detta knappast kunna vara fallet utan att de separeras. Det verkar inte finnas några belägg i litteraturen för parning av nattfjärilar i flygning. I de flesta Lepidoptera och i övervägande nattliga mikrolepidoptera (nattfjärilar) ”kallar” däremot honorna på hanarna med feromoner och bestämmer på så sätt parningsplatserna (Groot, 2014). De kan flyga före parningen, men i de flesta fall verkar de vara fastsittande och fläktande för feromonfördelning. Könen som ropar eller söker kan dock också vara omvända i vissa taxa, men för Tineidae rapporteras honan vara det ropande könet (Wagner och Rosovsky, 1991; Scoble, 1995). I vilket fall som helst var platsen för hartsflödet, dvs. biotopen med de bärnstensproducerande träden, troligen också livsmiljön för det klädsmal som beskrivs här.
KONKLUSION
En tineoid mal med en anmärkningsvärd hanlig genitalapparat beskrivs som den nya arten och släktet Forcepsites michalskii från eocen baltisk bärnsten. Honan och hanen är bevarade tillsammans i nära position och med buken orienterad mot varandra, vilket tyder på att dessa malar fastnade i det färska hartset under parningen. Ett starkt laminärt hartsflöde, vilket indikeras av flera drivna delar av honans kropp, kan ha dragit isär dem på det korta avstånd som nu observeras. Detta exemplar skulle därför kunna utgöra det första exemplet på fossila Lepidoptera som fångats i copula.
Det antas att Forcepsites michalskii var en skogslevande art, eftersom individer som fångats i copula endast kan dokumenteras från den plats där denna art levde och parade sig.
Hon och hane av F. michalskii uppvisar sexuell dimorfism i kroppsstorlek, vinglängd, proportioner på buken, storleken på de sammansatta ögonen och morfologin på maxillärpalperna, samt på antennerna. Baserat på de tydligt större och tjockare antennerna hos hanen samt på de större ögonen föreslås att honan skulle kunna vara det kallande könet hos denna art, vilket också rapporterats för existerande Tineidae-arter.
NACKMEDDELANDEN
A. Michalski (Wroclaw, Polen) tackas för att han tillhandahöll detta exceptionella exemplar och för att han ledde vår uppmärksamhet till det. M. Schwarz från Oberösterreichisches Landesmuseum (Linz, Österrike) gav vänligt stöd för genomgång av deras Lepidoptera-samling. MKH fick ett stipendium från Studienstiftung des deutschen Volkes och tackar S. Harzsch (Greifswald, Tyskland) för hans stöd. Vi tackar M. Heikkilä och C. Haug (München, Tyskland) för värdefulla vetenskapliga kommentarer till manuskriptet, liksom J. Krieger (Greifswald, Tyskland) för kommentarer och C. MacGraphery (Freising, Tyskland) och Y. Goldman (Collinsville, CT, USA) för att kontrollera den engelska texten som modersmålstalare. Dessutom tackar vi två anonyma granskare för deras användbara kommentarer.
Arillo, A. 2007. Paleoetologi: fossila beteenden i bärnsten. Geologica Acta, 5(2):159-166. https://doi.org/10.1344/105.000000301
Börner, C. 1925. Lepidoptera, Schmetterlinge, s. 358-387. I Brohmer, P. (red.). Fauna von Deutschland. Ein Bestimmungsbuch unserer heimischen Tierwelt. Quelle und Meyer, Leipzig.
Borkent, A. 1995. Biting Midges in the Cretaceous Amber of North America (Diptera: Ceratopogonidae). Backhuys Publishers, Leiden, Nederländerna.
Boucot, A.J. and Poinar Jr, G.O. 2010. Fossil Behavior Compendium. CRC Press, Boca Raton, s. 147-151.
Coty, D., Aria, C., Garrouste, R., Wils, P., Legendre, F. och Nel, A. 2014. Den första syninklusionen av myrtermiter i bärnsten med CT-scan-analys av taphonomin. PLOS one, 9(8):e104410. https://doi.org/10.1371/journal.pone.0104410
Deering, M.D. och Scriber, J.M. 2002. Bioassays i fält visar att det finns en asymmetri i de heterospecifika parningspreferenserna mellan hybridiserande nordamerikanska Papilio-fjärilsarter (Lepidoptera: Papilionidae). Journal of Ethology 20:25-33. https://doi.org/10.1007/s10164-002-0050-2
Fletcher, T.B. 1940. Nya generiska namn för microlepidoptera. Entomologist’s Record, 52:17-19.
Ganyard, M.C. and Brady, Jr.U.E. 1972. Interspecifik attraktion hos Lepidoptera i fält. Annals of the Entomological Society of America, 65 (6): 1279-1282. https://doi.org/10.1093/aesa/65.6.1279
Gröhn, C. 2016. Inkluderingar i baltisk bärnsten. Wachholtz Verlag – Murmann Publishers, Kiel, Hamburg.
Groot, A.T. 2014. Cirkadiska rytmer för sexuella aktiviteter hos nattfjärilar: En genomgång. Frontiers in Ecology and Evolution 2:43. https://doi.org/10.3389/fevo.2014.00043
Hoffeins, C. och Hoffeins, H.W. 2003. Studier av förekomsten av inneslutningar i bärnsten från Baltikum och Bitterfeld (tertiär, eocen) från icke-selekterade samlingar med särskild hänsyn till ordningen Diptera. Studia Dipterologica, 10(2):381-392.
Kozlov, M.V. 1987. Nya malliknande Lepidoptera från baltisk bärnsten. Paleontological Journal, 4:59-67.
Kozlov, M.V. 1988. Paleontologi av lepidopteraner och problem med fylogenin i ordningen Papilionida, s. 16-69. I Ponomarenko A.G. (red.), The Mesozoic-Cenozoic Crisis in the Evolution of Insects. Academy of Sciences, Moscow.
Krzeminska, E., Krzeminski, W., Haenni, J.-P. och Dufour, C. 1991. Les Fantomes De L’Ambre. Muséed’histoire naturelle de Neuchâtel, Neuchâtel, Schweiz.
Kusnezov, N. 1941. En revidering av Amber Lepidoptera. Paleontologiska institutet, USSR Academy of Sciences, Moskva & Leningrad.
Latreille, P.A. 1810. Allmänna överväganden om den naturliga ordningen hos de djur som ingår i klasserna kräftdjur, spindeldjur och insekter: med en metodisk tabell över deras släkten, ordnade i familjer. Paris, Schoell.
Menken, S.B.J., Boomsma, J.J. och van Nieukerken, E.J. 2009. Storskaliga evolutionära mönster av värdväxtföreningar hos Lepidoptera. Evolution, 64(4):1098-1119. https://doi.org/10.1111/j.1558-5646.2009.00889.x
Rebel, H. 1934. Bernstein-Lepidopteren (aus der Sammlung Bachofen-Echt). Palaeobiologica, 6:1-16.
Rebel, H. 1935. Bernstein-Lepidopteren. Entomologische Zeitschrift Iris, 49:162-186.
Regier, J.C., Mitter, C., Davis, D.R., Harrison, T.L., Sohn J.-C., Cummings, M.P., Zwick, A., and Mitter, K.T. 2014. En molekylär fylogeni och reviderad klassificering för de äldsta ditrysia-motledslinjerna (Lepidoptera: Tineoidea), med implikationer för förfädernas matvanor hos den megadiversifierade Ditrysia. Systematic Entomology, 40(2). https://doi.org/10.1111/syen.12110
Scoble, M.J. 1995. Lepidoptera – form, funktion och mångfald. The Natural History Museum / Oxford University Press, Oxford.
Skalski, A.W. 1973. Studier av Lepidoptera från fossila hartser. Del II. Epiborkhausenites obscurotrimaculatus gen. et sp. nov. (Oecophoridae) och en Tineid-mot som upptäckts i baltisk bärnsten. Acta Palaeontologica Polonica, 18(1):153-160.
Skalski, A.W. 1974. Zwei neue Gattungen und Arten der Familie Tineidae aus dem Baltischen Bernstein. Studien an Lepidopteren aus fossilen Harzen V. Beiträge zur Entomologie, 1/4(24):97-104.
Skalski, A.W. 1977. Studier av Lepidoptera från fossila hartser. Del I. Allmänna anmärkningar och beskrivningar av nya släkten och arter i familjerna Tineidae och Oecophoridae från baltiskt bärnsten. Prace Museum Ziemi, 26:3-24.
Sohn, J.-C., Labandeira, C., Davis, D. och Mitter, C. 2012. En kommenterad katalog över fossila och subfossila Lepidoptera (Insecta: Holometabola) i världen. Zootaxa, 3286:1-132.
Sohn, J.-C. and Lamas, G. 2013. Korrigeringar, tillägg och nomenklaturanmärkningar till den nyligen publicerade världskatalogen över fossila och subfossila Lepidoptera. Zootaxa, 3599:395-399.
Sontag, E. 2003. Djurinklusioner i ett prov av oselekterad baltisk bärnsten. Acta Zoologica Cracoviensia, 46 (suppl.-Fossil Insects):431-440.
van Nieukerken, E.J., Kaila, L., Kitching, I.J., Kristensen, N.P., Lees, D.C., Minet, J., Mitter, C., Mutanen, M., Regier, J.C., Simonsen, T.J., Wahlberg, N., Yen, S.-H., Zahiri, R., Adamski, D., Baixeras, J., Bartsch, D., Bengtsson, B.Å., Brown, J.W., Bucheli, S.R., Davis, D.R., De Prins, J., De Prins, W., Epstein, M.E., Gentili-Poole, P., Gielis, C., Hätteschwiler, P., Hausmann, A., Holloway, J.D., Kallies, A., Karsholt, O., Kawahara, A.Y., Koster, S.J.C., Kozlov, M.V., Lafontaine, J.D., Lamas, G., Landry, J.-F., Lee, S., Nuss, M., Park, K.-T., Penz, C., Rota, J., Schintlmeister, A., Schmidt, B.C., Sohn, J.-C., Solis, M.A., Tarmann, G.M., Warren, A.D., Weller, S., Yakovlev, R.V., Zolotuhin, V.V. och Zwick, A. 2011. Ordningen Lepidoptera Linnaeus, 1758. I Zhang, Z.-Q. (red.), Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa, 3148:212-221.
von Linné, C. 1758. översatt av William Turton (1806). Volym 1. A General System of Nature: Through the Three Grand Kingdoms of Animals, Vegetables, and Minerals, Systematically Divided into their Several Classes, Orders, Genera, Species, and Varieties. Lackington, Allen, and Co, London.
Wagner, D.L. och Rosovsky, J. 1991. Parningssystem hos primitiva Lepidoptera, med tonvikt på det reproduktiva beteendet hos Korscheltellus gracilis (Hepialidae). Zoological Journal of the Linnean Society, 102:277-303. https://doi.org/10.1111/j.1096-3642.1991.tb00003.x
Weitschat, W. och Wichard, W. 2002. Atlas över växter och djur i baltisk bärnsten. Verlag Friedrich Pfeil, München.
Weitschat, W. 2009. Jäger, Gejagte, Parasiten und blinde Passagiere – Momentaufnahmen aus dem Bernsteinwald. Denisia, 26(86):243-256.
Wichard, W., Gröhn, C., and Seredszus, F. 2009. Vattenlevande insekter i baltisk bärnsten. Verlag Kessel, Remagen-Oberwinter.