- Molii de împerechere (Tineidae, Ditrysia, Lepidoptera) conservate ca incluziune comportamentală congelată în chihlimbarul baltic (Eocen)
- ABSTRACT
- INTRODUCERE
- MATERIALE ȘI METODE
- PALEONTOLOGIE SISTEMATICĂ
- DISCUȚII
- Coocurență aleatorie sau o împerechere păstrată?
- Tafonomie
- Dimorfism sexual
- Habitat
- CONCLUZIE
- ACKNOWLEDGMENTS
Molii de împerechere (Tineidae, Ditrysia, Lepidoptera) conservate ca incluziune comportamentală congelată în chihlimbarul baltic (Eocen)
Thilo C. Fischer și Marie K. Hörnig
Numărul articolului: 22.1.7
https://doi.org/10.26879/829
Copyright Paleontological Society, February 2019
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 22 Octobere 2017. Acceptare: 23 ianuarie 2019
{flike id=2421}
ABSTRACT
Investigarea incluziunilor de chihlimbar oferă o perspectivă unică asupra biologiei animalelor care au trăit cu milioane de ani în urmă, datorită conservării lor excepționale în condiții aproape asemănătoare vieții. Acest lucru nu este valabil numai pentru aspectele morfologice, aspecte ale comportamentului pot fi, de asemenea, deduse din aceste fosile.
În acest caz, Microlepidoptera este raportată din chihlimbarul baltic din Eocen, cel mai probabil ca o primă apariție de Lepidoptera fosilă găsită în copula. Moliile aparțin grupului Tineidae (moliile tineide includ moliile de haine) și sunt descrise ca o nouă specie și un nou gen, Forcepsites michalskii. Organul de încleștare al masculului este destul de neobișnuit pentru moliile din chihlimbarul baltic, cu o parte ventrală și una dorsală, fiecare terminată în două protuberanțe paralele, asemănătoare unor spini, și care se pare că nu este formată de valvae. Prezența comună a masculului și a femelei, cel mai probabil în copula, a permis studierea directă a dimorfismului lor sexual și există dovezi că habitatul acestei specii este locul de producere a rășinii în pădurea de chihlimbar baltic. Din dimorfismul sexual al antenelor și morfologia ochilor se interpretează că femela a fost sexul de apel pentru împerechere la această specie.
Thilo C. Fischer. Förderverein der Bayerischen Staatssammlung für Paläontologie und Historische Geologie München Richard-Wagner-Straße 10, D-80333 München, Germania.
Marie K. Hörnig. Universitatea din Greifswald, Institutul și Muzeul de Zoologie, Citologie și Biologie Evolutivă, Soldmannstr. 23, D-17489 Greifswald, Germania.
CUVINTE CHEIE: Clothes moths; habitat în copula; Microlepidoptera; specii și genuri noi; dimorfism sexual
Citire finală: Fischer, Thilo C, și Hörnig, Marie K. 2019. Molii de împerechere (Tineidae, Ditrysia, Lepidoptera) conservate ca incluziune de comportament congelat în chihlimbarul baltic (Eocen). Palaeontologia Electronica 22.1.7A 1-11. https://doi.org/10.26879/829
palaeo-electronica.org/content/2019/2421-mating-moths
Copyright: Februarie 2019 Paleontological Society.
Acest articol este un articol cu acces liber distribuit în conformitate cu termenii Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), care permite utilizatorilor să copieze și să redistribuie materialul în orice mediu sau format, cu condiția să nu fie folosit în scopuri comerciale și să fie citat autorul original și sursa, cu indicații în cazul în care se fac modificări.
creativecommons.org/licenses/by-nc-sa/4.0/
http://zoobank.org/BCF64D89-6879-4805-8021-7CE926CE4B12
INTRODUCERE
Insectele care se împerechează sunt rareori întâlnite în chihlimbar ca și cazuri ale așa-numitului „comportament înghețat” (Arillo, 2007, și referințe în acest sens). Astfel de insecte au fost raportate din chihlimbarul baltic din Eocen, de exemplu din grupurile Diptera, Coleoptera, Hemiptera, Hymenoptera și destul de rar Trichoptera (Weitschat și Wichard, 2002; Weitschat, 2009; Boucot și Poinar Jr., 2010; Gröhn, 2016).
În chihlimbarul baltic, Lepidoptera sunt în general rare și apar la aproximativ una din 250 de incluziuni (pentru comparație: Dipterele reprezintă aproximativ 64% din toate insectele din chihlimbarul baltic, Lepidoptera reprezintă aproximativ 0,4 %; Hoffeins și Hoffeins 2003; Gröhn 2016). Ca și în cazul altor insecte, imagos conservate în copula sunt mult mai rare, iar Lepidoptera în copula nu au fost încă descrise, nici măcar în alte ambre sau din localități de insecte din afara ambrei (Sohn et al., 2012; Sohn și Lamas, 2013). O apariție in copula permite afilierea de bună credință a ambelor forme sexuale una față de cealaltă și a ambelor împreună la o specie distinctă și, prin urmare, descrierea exactă a dimorfismului sexual al unei specii. În mod alternativ, dimorfismul sexual din insectele fosile poate fi studiat prin mijloace statistice doar atunci când descoperirile unei specii sunt abundente, așa cum este cazul multor Nematocera (de ex, Borkent, 1995).
Tineidae sunt reprezentanți bazali foarte diverși ai grupului de lepidoptere Ditrysia (Regier et al., 2014), larvele lor se hrănesc cu o gamă largă de substraturi (detritus, ciuperci, licheni sau cheratină) (Scoble, 1995), fiind astfel generaliste (85,7%) (Menken et al., 2009). Ca incluziuni în chihlimbarul baltic, Tineidae reprezintă aproximativ un sfert din toate descoperirile de lepidoptere (Skalski, 1977). Aici, este descrisă o specie nouă și rară de Tineidae cu formele sale sexuale dintr-o prezență putativă în copula.
MATERIALE ȘI METODE
Specimenul de chihlimbar a fost găsit și achiziționat de la Artur Michalski (Wroclaw, Polonia). A fost conservat prin tratare cu un lac acrilic și provine din colecția autorului (TF) cu numărul de colecție 7131. Depozitarea a avut loc la temperatură constantă în saci de plastic cu clips în interiorul unor cutii metalice, cu excepția oxigenului și a luminii. Va fi depozitat la Bayerische Paläontologische Staatssammlung München cu numărul SNSB-BSPG 2017 I 113.
Exemplarul a fost fotografiat cu un Canon EOS 70d echipat cu un obiectiv Canon MP-E 65 mm și un Canon Twin Flash MT-24. Pentru a evita reflexiile, filtrele de polarizare au fost montate și ajustate în fața blițurilor și a obiectivului camerei (lumină polarizată încrucișată). Pentru a reduce convexitatea și concavitățile suprafeței chihlimbarului, s-a aplicat o picătură de glicerină și s-a acoperit cu un film de acoperire. Stivele z de imagini înregistrate au fost fuzionate cu Combine ZP și prelucrate ulterior cu Adobe Photoshop CS 3. Incluzia nu a putut fi studiată prin microtomografie computerizată, din cauza prezenței incluziunilor metalice, aparent piritice, care provoacă artefacte puternice.
PALEONTOLOGIE SISTEMATICĂ
Sistematică (conform van Nieukerken et al., 2011)
Ordine LEPIDOPTERA Linnaeus, 1758
Clasa DITRYSIA Börner, 1925
Superfamilia TINEOIDEA Latreille, 1810
Familia TINEIDAE Latreille, 1810
Genul Forcepsites nov. gen.
zoobank.org/DDB6884D-845A-4A87-981E-A2953ADD8964
Etymology. „Forceps”, forceps în latină, referindu-se la organul de încleștare al masculului.
Specie Forcepsites michalskii nov. spec.
zoobank.org/5494D58E-7773-473C-AFB2-140DDD049226
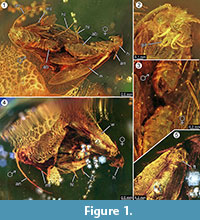 Holotypus. Specimen ex coll. Fischer nr. 7131, Figura 1.2
Holotypus. Specimen ex coll. Fischer nr. 7131, Figura 1.2
Locus typicus. Mina de chihlimbar din Yantarni, GUS
Stratum typicum. „Blaue Erde” (Eocen superior – Oligocen inferior)
Etimologie. Numele speciei michalskii este ales pentru a-l recunoaște pe Artur Michalski pentru furnizarea acestui specimen.
Depozit. Bayerische Staatssammlung für Paläontologie und Geologie, München, Germania; număr de acces SNSB-BSPG 2017 I 113.
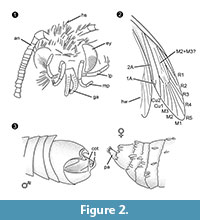
Diagnosticarea genului. Gen monotipic, având ca caracter definitoriu organul de încleștare al masculului (figura 2.3) cu o parte ventrală și una dorsală, fiecare terminată cu două protuberanțe paralele, asemănătoare cu spinii.
 Diagnoza speciei. Caracterele calitative și cantitative respective ale imagoanelor feminine și masculine (figura 1) au fost determinate și sunt rezumate în tabelul 1.
Diagnoza speciei. Caracterele calitative și cantitative respective ale imagoanelor feminine și masculine (figura 1) au fost determinate și sunt rezumate în tabelul 1.
Diagnostic diferențial. Combinația de caractere proboscis mic cu galee separate, palpezi maxilari lungi și pliați cu cinci segmente, palpezi labiali îndreptați în sus și ușor îndoiți cu peri laterali, cap cu multe solzi erecți asemănători părului sugerează puternic o afiliere la Tineidae, dar niciunul dintre acestea nu este apomorfic, și nici nu este cunoscută în prezent o astfel de apomorfie pentru Tineidae adulte (Regier et al., 2014). Venele R4 și R5 nu sunt fuzionate / stâlcite, ceea ce ar fi indicativ pentru Gelechioidea (Gelechiidae și Oecophoridae) (Scoble, 1995), celălalt grup major de lepidoptere găsit în chihlimbarul baltic. Specia nu seamănă cu niciun reprezentant descris al Tineidae sau al altor familii din chihlimbarul baltic (Rebel, 1934, 1935; Kuznezov, 1941; Skalski, 1973; Kozlov, 1987, 1988; Sohn et al., 2012; Sohn și Lamas, 2013), în special în ceea ce privește aparatul genital masculin. Cel mai asemănător pare Tineosemopsis decurtatus Skalski 1974 (femelă cunoscută), dar se deosebește prin absența firelor de păr erecte la vertex și prin ochii săi mai mici. Tillyardinea eocaenica Kusnezov 1941 (femelă), care are solzi erecți la vertex și o venație similară, diferă prin frons și palpi labiali. Simulotinea intermedia Skalski 1977 are un habitus asemănător, dar se deosebește și prin prezența unei vene bifurcate, care împarte celula mare din aripa anterioară (M2+3).
Tafonomie. Masculul și femela lui Forcepsites michalskii sunt fosilizate în apropiere și cu partea posterioară a abdomenului îndreptată unul spre celălalt. Cele două incluziuni se află într-o piesă cilindrică de chihlimbar baltic neautoclavizat, cu un diametru de 15 mm și o înălțime de 11 mm, care este împărțită la aproximativ jumătate din înălțime de o margine neuniformă între două straturi de curgere a rășinii. Această margine prezintă un strat de pirita ca parte a planului său. Unul dintre fluxurile de rășină conține cele două molii care își expun părțile dorsale respective la această margine. Se păstrează solzi lungi care derivă din marginea aripii femelei. Imago femelă a pierdut două picioare, iar toracele este deteriorat. Celălalt flux conține serii neliniare de bule. Ambii indivizi par nou apăruți, după cum reiese din veșmântul complet de solzi de pe aripi și din faptul că aripile nu sunt zdrențuite. Nu au putut fi identificate organe olfactive pe aripi, abdomen sau picioare. Singurele sinincluziuni păstrate cu microlepidopterele sunt tricomii, tipici pentru chihlimbarul baltic.
DISCUȚII
Coocurență aleatorie sau o împerechere păstrată?
Moliile închise sunt reprezentanți masculi și femele ai aceleiași specii și sunt dispuse în apropiere, cu abdomenele orientate unul spre celălalt. Cu toate acestea, organele genitale ale moliilor nu sunt în contact direct, ci într-o poziție apropiată. Orientarea dorsală/ventrală a masculului și a femelei diferă ușor, ceea ce este cauzat de o ușoară rotație în cadrul axei corpului (în special a masculului). Orientarea părții posterioare a abdomenului ambelor molii este una spre cealaltă.
Sunt cunoscute mai multe cazuri în care situația de închidere în rășină încă lichidă poate induce un răspuns de stres, cum ar fi ovipoziția (Weitschat, 2009). Inducerea comportamentului de împerechere de către fluxul de rășină nu este, în general, plauzibilă, dar situația de închidere demonstrează că au fost posibile ușoare mișcări în rășina proaspătă.
Există, de asemenea, întrebarea dacă această constatare ar putea reprezenta, de asemenea, o co-ocurență aleatorie. Probabilitatea pentru o astfel de co-ocurență aleatorie poate fi estimată în mod aproximativ. Printre incluziunile de insecte, moliile apar cu o frecvență de aproximativ una din 250 de incluziuni în chihlimbarul baltic (Krzeminskaet al., 1991 (0,6%); Hoffeins și Hoffeins, 2003 (0,4%); Sontag, 2003 (0,27%), și observația proprie). Având în vedere un chihlimbar individual cu o molie inclusă, șansa de apariție aleatorie a unei alte molii este pur și simplu de 1:250 (moliile nu apar în roiuri). Coocurența aceluiași taxon, sau a unuia abundent care ar putea fi confundat, este estimată la 1:10 în cel mai bun caz (Skalski, 1977), și încă aproximativ 1:2 pentru a avea sexe diferite. Luând în considerare șansele suplimentare de aranjament apropiat, atât alinierea axelor corpului, cât și o orientare adecvată a corpului, o probabilitate pentru o co-ocurență aleatorie care ar putea fi interpretată greșit ca un eveniment de împerechere este mult mai mică decât frecvența observată ca fiind una din 920 de incluziuni de lepidoptere în colecția autorului studiat.
În concluzie, cea mai probabilă interpretare pentru această constatare este că moliile se aflau în copulație, iar organele genitale au fost în contact, dar indivizii au fost ușor distrași unul de celălalt din cauza fluxului de rășină (a se vedea discuția de mai jos).
Tafonomie
Relevanța analizei atente a fluxurilor de rășină pentru tafonomia incluziunilor de chihlimbar a fost demonstrată de Coty et al. (2014) cu studiul lor privind o interacțiune furnică – termită păstrată în chihlimbarul mexican din Chiapas.
Incidența unui eveniment in copula este puternic susținută de descoperirea unor solzi lungi derivați de pe marginea aripii femelei, care au derapat pe o anumită distanță în direcția masculului. Acest lucru ar putea indica un flux laminar puternic de rășină în acest loc, care s-a deplasat de-a lungul axei lor comune și care ar fi putut să le îndepărteze pe distanța scurtă observată acum. În timpul împerecherii moliilor, masculul poate pierde contactul cu locul de împerechere și poate fi fixat de femelă doar prin organul de încleștare, doar femela făcând contact cu suprafața. Un astfel de comportament și tragerea din cauza fluxului brusc de rășină ar putea fi motivul pentru care femela și-a pierdut ambele picioare posterioare. Pe partea dorsală a moliilor, care a fost expusă la suprafața convexă a acestui prim flux laminar de rășină, aripa femelei și toracele de sub ea sunt parțial deteriorate (figura 1.5). Se presupune că aceste zone distruse reprezintă urme de hrănire cauzate de alte insecte, cel mai probabil furnici, așa cum se observă adesea în exemplarele de chihlimbar. Prin urmare, după primul flux de rășină ca eveniment de încorporare, părțile dorsale vor fi fost expuse pentru o perioadă de timp, suficient de lungă pentru a permite prădătorilor să pășească pe această suprafață solidă. Al doilea flux de rășină, care a acoperit această parte mai târziu, ar fi putut fi un flux rapid și turbulent de rășină, după cum indică o serie de bule mici dispuse în curbe neregulate găsite în interiorul acestuia. Acesta ar fi un proces tafonomic tipic pentru un așa-numit „Schlaube” de chihlimbar baltic (Gröhn, 2016).
Dimorfism sexual
Dimorfismul sexual este dificil de observat la speciile fosile, de la care se cunosc, de obicei, doar câteva exemplare. Caracterele care nu sunt influențate de sex trebuie să fie diagnostice pentru a permite o determinare sigură a speciilor ca bază pentru diferențierea ulterioară între sexe și elucidarea dimorfismului sexual. Având în vedere speciile existente cu un dimorfism sexual ridicat, de exemplu, cu femele brachypterous, cum este cazul multor Psychidae (Scoble, 1995), nu ar fi posibil să se deducă că ambele sexe aparțin unei singure specii. Prin urmare, numai cazurile de dimorfism sexual slab pot fi recunoscute în mod fiabil la speciile fosile prin mijloace statistice și fără a le găsi în copula. Acest lucru subliniază importanța comportamentului de împerechere identificat la speciile fosile pentru descrierea unor astfel de cazuri de dimorfism sexual puternic. Astfel de constatări conservate de împerechere fosilă pot aproba, de asemenea, corelarea formelor dimorfice sexuale cunoscute anterior pentru a aparține unei singure specii, chiar și fără posibilitatea unui sprijin statistic prea mare. Există o anumită șansă ca un eveniment de împerechere conservat să reprezinte o împerechere interspecifică sau intergenerică, așa cum sunt cunoscute la molii și fluturi (Ganyard și Brady, 1972; Deering și Scriber, 2002). Cu toate acestea, aceste fenomene sunt mult mai puțin abundente decât împerecherea intraspecifică și este puțin probabil să fie găsite ca o incluziune fosilă rară.
Un dimorfism sexual non-allometric observat la Forcepsites michalskii este reprezentat de galeelele separate în probosciscul la mascul. Acest lucru poate face aluzie la o stare nefuncțională, imagosul mascul al acestei specii fiind nealimentar, așa cum se observă la multe Psychidae (masculi) existente cu proboscis vestigială sau absentă (Scoble, 1995). Dimorfismele sexuale alometrice găsite aici sunt: (1) Femela este mai mare decât masculul, atât în ceea ce privește dimensiunea corpului, cât și lungimea aripilor. Cu toate acestea, raporturile respective sunt aproape identice. (2) La femelă, abdomenul este relativ și absolut mai lat în comparație cu cel al masculului. (3) Antenele masculului sunt mult mai mari și mai groase decât la femela imago. La mai multe specii existente antenele sunt dimorfice din punct de vedere sexual (Scoble, 1995), antenele masculine bine dezvoltate servesc la chimiosensibilizarea feromonilor feminini. (4) Palpii maxilari ai masculului sunt de două ori mai groși decât la femelă și au o structură cu nervuri. (5) Ochii masculului sunt, de asemenea, mai mari, dar numărul de ommatidii este aproximativ același. Ochii mai mari vor primi doar mai multă lumină și ar putea fi mai sensibili; cu toate acestea, rezoluția ar fi aceeași.
Recunoașterea pe distanțe lungi a moliilor femele care cheamă are loc prin intermediul chemoreceptorilor masculi, în timp ce recunoașterea pe distanțe scurte se bazează în schimb pe identificarea vizuală (Groot, 2014). La Forcepsites michalskii, se pare, de asemenea, că femela a fost sexul apelant – după cum sugerează aparatul antenar și ochii mai mari ai masculului.
Habitat
Pe baza discuției prezentate mai sus, este clar că indivizii prinși în copula pot apărea numai în locul în care această specie a trăit și s-a împerecheat. Indivizii singuri pot fi plutiți în derivă de vânt sau pot vizita locuri fără habitat, cum ar fi insectele acvatice rar întâlnite în chihlimbarul baltic (Wichardet al., 2009); în schimb, pentru indivizii care se împerechează, acest lucru ar putea cu greu să se întâmple fără a fi separați. Se pare că nu există dovezi în literatura de specialitate privind împerecherea moliilor în zbor. În schimb, la majoritatea lepidopterelor și la microlepidopterele predominant nocturne (molii), femelele „cheamă” masculii prin feromoni, determinând astfel locurile de împerechere (Groot, 2014). Ele pot zbura înainte de împerechere, dar în cele mai multe cazuri par să fie sesile și să facă evantai pentru distribuirea feromonilor. Cu toate acestea, sexele care cheamă sau caută pot fi, de asemenea, inversate în unii taxoni, dar pentru Tineidae se raportează că femela este sexul care cheamă (Wagner și Rosovsky, 1991; Scoble, 1995). În orice caz, locul de curgere a rășinii, adică biotopul cu arbori producători de chihlimbar, cel mai probabil a fost, de asemenea, habitatul moliei de haine descrisă aici.
CONCLUZIE
Se descrie o molie tineoidă cu un aparat genital masculin remarcabil, noua specie și genul Forcepsites michalskii din chihlimbarul baltic eocen. Femela și masculul sunt conservate împreună în poziție apropiată și cu abdomenul orientat unul spre celălalt, sugerând că aceste molii au fost prinse în rășină proaspătă în timpul împerecherii. Un flux laminar puternic de rășină, care este indicat de mai multe părți ale corpului femelei, ar fi putut să îi îndepărteze pe cei doi pe distanța scurtă observată acum. Acest specimen ar putea, prin urmare, să reprezinte primul exemplu de Lepidoptera fosilă capturată în copula.
Se presupune că Forcepsites michalskii a fost o specie care locuia în pădure, deoarece indivizii capturați în copula pot fi documentați doar din locul în care această specie a trăit și s-a împerecheat.
Femele și masculii de F. michalskii prezintă dimorfisme sexuale în ceea ce privește mărimea corpului, lungimea aripilor, proporția abdomenului, mărimea ochilor compuși și morfologia palpilor maxilari, precum și a antenelor. Pe baza antenelor distincte mai mari și mai groase ale masculului, precum și a ochilor mai mari, se sugerează că femela ar putea fi sexul de apel la această specie, așa cum s-a raportat și pentru speciile de Tineidae existente.
ACKNOWLEDGMENTS
A. Michalski (Wroclaw, Polonia) este recunoscut pentru că a furnizat acest specimen excepțional și ne-a atras atenția asupra lui. M. Schwarz de la Oberösterreichisches Landesmuseum (Linz, Austria) a sprijinit cu amabilitate examinarea colecției lor de lepidoptere. MKH a fost finanțat cu amabilitate de o bursă de la Studienstiftung des deutschen Volkes și îi mulțumește lui S. Harzsch (Greifswald, Germania) pentru sprijin. Recunoaștem comentariile științifice valoroase pe care M. Heikkilä și C. Haug (München, Germania) le-au făcut asupra manuscrisului, precum și comentariile lui J. Krieger (Greifswald, Germania) și sprijinul acordat de C. MacGraphery (Freising, Germania) și Y. Goldman (Collinsville, CT, SUA) pentru verificarea textului în limba engleză, în calitate de vorbitori nativi. În plus, mulțumim celor doi recenzenți anonimi pentru comentariile lor utile.
Arillo, A. 2007. Paleoetologie: comportamente fosilizate în chihlimbar. Geologica Acta, 5(2):159-166. https://doi.org/10.1344/105.000000301
Börner, C. 1925. Lepidoptera, Schmetterlinge, p. 358-387. În Brohmer, P. (ed.). Fauna von Deutschland. Ein Bestimmungsbuch unserer heimischen Tierwelt. Quelle und Meyer, Leipzig.
Borkent, A. 1995. Biting Midges in the Cretaceous Amber of North America (Diptera: Ceratopogonidae). Backhuys Publishers, Leiden, Țările de Jos.
Boucot, A.J. și Poinar Jr, G.O. 2010. Fossil Behavior Compendium. CRC Press, Boca Raton, Boca Raton, p. 147-151.
Coty, D., Aria, C., Garrouste, R., Wils, P., Legendre, F. și Nel, A. 2014. Prima sinincluzie de furnici-termite în chihlimbar cu analiză CT-scan a tafonomiei. PLOS one, 9(8):e104410. https://doi.org/10.1371/journal.pone.0104410
Deering, M.D. și Scriber, J.M. 2002. Testele biologice pe teren arată asimetria preferințelor de împerechere heterospecifică între speciile de fluturi Papilio hibridizanți din America de Nord (Lepidoptera: Papilionidae). Journal of Ethology 20:25-33. https://doi.org/10.1007/s10164-002-0050-2
Fletcher, T.B. 1940. Noi nume generice pentru microlepidoptere. Entomologist’s Record, 52:17-19.
Ganyard, M.C. și Brady, Jr.U.E. 1972. Atracția interspecifică la Lepidoptera în câmp. Analele Societății Entomologice a Americii, 65 (6): 1279-1282. https://doi.org/10.1093/aesa/65.6.1279
Gröhn, C. 2016. Incluziuni în chihlimbarul baltic. Wachholtz Verlag – Murmann Publishers, Kiel, Hamburg.
Groot, A.T. 2014. Ritmurile circadiene ale activităților sexuale la molii: O analiză. Frontiere în ecologie și evoluție 2:43. https://doi.org/10.3389/fevo.2014.00043
Hoffeins, C. și Hoffeins, H.W. 2003. Studii privind abundența incluziunilor în chihlimbarul Baltic și Bitterfeld (Terțiar, Eocen) din colecții neselectate, cu referire specială la ordinul Diptera. Studia Dipterologica, 10(2):381-392.
Kozlov, M.V. 1987. Noi lepidoptere asemănătoare moliilor din chihlimbarul baltic. Paleontological Journal, 4:59-67.
Kozlov, M.V. 1988. Paleontologia lepidopterelor și probleme de filogenie a ordinului Papilionida, p. 16-69. În Ponomarenko A.G. (ed.), The Mesozoic-Cenozoic Crisis in the Evolution of Insects. Academia de Științe, Moscova.
Krzeminska, E., Krzeminski, W., Haenni, J.-P., și Dufour, C. 1991. Les Fantomes De L’Ambre. Muséed’histoire naturelle de Neuchâtel, Neuchâtel, Elveția.
Kusnezov, N. 1941. O revizuire a lepidopterelor de chihlimbar. Institutul de Paleontologie, Academia de Științe a URSS, Moscova & Leningrad.
Latreille, P.A. 1810. Considerații generale asupra ordinii naturale a animalelor care alcătuiesc clasele de crustacee, arahnide și insecte: cu un tabel metodic al genurilor lor, aranjate pe familii. Paris, Schoell.
Menken, S.B.J., Boomsma, J.J. și van Nieukerken, E.J. 2009. Modele evolutive pe scară largă ale asociațiilor cu plante gazdă la Lepidoptera. Evolution, 64(4):1098-1119. https://doi.org/10.1111/j.1558-5646.2009.00889.x
Rebel, H. 1934. Bernstein-Lepidopteren (aus der Sammlung Bachofen-Echt). Palaeobiologica, 6:1-16.
Rebel, H. 1935. Bernstein-Lepidopteren. Entomologische Zeitschrift Iris, 49:162-186.
Regier, J.C., Mitter, C., Davis, D.R., Harrison, T.L., Sohn J.-C., Cummings, M.P., Zwick, A., și Mitter, K.T. 2014. O filogenie moleculară și o clasificare revizuită pentru cele mai vechi linii de molii ditrysia (Lepidoptera: Tineoidea), cu implicații pentru obiceiurile ancestrale de hrănire ale mega-diversei Ditrysia. Systematic Entomology, 40(2). https://doi.org/10.1111/syen.12110
Scoble, M.J. 1995. Lepidoptera – Formă, funcție și diversitate. The Natural History Museum / Oxford University Press, Oxford.
Skalski, A.W. 1973. Studies on the Lepidoptera from fossil resins. Part II. Epiborkhausenites obscurotrimaculatus gen. et sp. nov. (Oecophoridae) și un Tineid-moth descoperit în chihlimbarul baltic. Acta Palaeontologica Polonica, 18(1):153-160.
Skalski, A.W. 1974. Zwei neue Gattungen und Arten der Familie Tineidae aus dem Baltischen Bernstein. Studien an Lepidopteren aus fossilen Harzen V. Beiträge zur Entomologie, 1/4(24):97-104.
Skalski, A.W. 1977. Studies on the Lepidoptera from fossil resins. Part I. General remarks and descriptions of new genres and species of the families Tineidae and Oecophoridae from Baltic amber. Prace Museum Ziemi, 26:3-24.
Sohn, J.-C., Labandeira, C., Davis, D., and Mitter, C. 2012. An annotated catalog of fossil and subfossil Lepidoptera (Insecta: Holometabola) of the world. Zootaxa, 3286:1-132.
Sohn, J.-C. și Lamas, G. 2013. Corecții, adăugiri și note nomenclaturale la catalogul mondial recent publicat de Lepidoptera fosilă și subfossilă. Zootaxa, 3599:395-399.
Sontag, E. 2003. Incluziuni animale într-un eșantion de chihlimbar baltic neselectat. Acta Zoologica Cracoviensia, 46 (suppl.-Fossil Insects):431-440.
van Nieukerken, E.J., Kaila, L., Kitching, I.J., Kristensen, N.P., Lees, D.C., Minet, J., Mitter, C., Mutanen, M., Regier, J.C., Simonsen, T.J., Wahlberg, N., Yen, S.-H., Zahiri, R., Adamski, D., Baixeras, J., Bartsch, D., Bengtsson, B.Å., Brown, J.W., Bucheli, S.R., Davis, D.R., De Prins, J., De Prins, W., Epstein, M.E., Gentili-Poole, P., Gielis, C., Hätteschwiler, P., Hausmann, A., Holloway, J.D., Kallies, A., Karsholt, O., Kawahara, A.Y., Koster, S.J.C., Kozlov, M.V., Lafontaine, J.D., Lamas, G., Landry, J.-F., Lee, S., Nuss, M., Park, K.-T., Penz, C., Rota, J., Schintlmeister, A., Schmidt, B.C., Sohn, J.-C, Solis, M.A., Tarmann, G.M., Warren, A.D., Weller, S., Yakovlev, R.V., Zolotuhin, V.V., și Zwick, A. 2011. Ordinul Lepidoptera Linnaeus, 1758. În Zhang, Z.-Q. (ed.), Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa, 3148:212-221.
von Linné, C. 1758. tradus de William Turton (1806). Volumul 1. Un sistem general al naturii: Prin cele trei mari regnuri ale animalelor, legumelor și mineralelor, împărțite sistematic în diferitele lor clase, ordine, genuri, specii și varietăți. Lackington, Allen, and Co, London.
Wagner, D.L. și Rosovsky, J. 1991. Sisteme de împerechere la lepidopterele primitive, cu accent pe comportamentul reproductiv al lui Korscheltellus gracilis (Hepialidae). Zoological Journal of the Linnean Society, 102:277-303. https://doi.org/10.1111/j.1096-3642.1991.tb00003.x
Weitschat, W. și Wichard, W. 2002. Atlas de plante și animale din chihlimbarul baltic. Verlag Friedrich Pfeil, München.
Weitschat, W. 2009. Jäger, Gejagte, Parasiten und blinde Passagiere – Momentaufnahmen aus dem Bernsteinwald. Denisia, 26(86):243-256.
Wichard, W., Gröhn, C., and Seredszus, F. 2009. Insecte acvatice în chihlimbarul baltic. Verlag Kessel, Remagen-Oberwinter.
.