- Parende nachtvlinders (Tineidae, Ditrysia, Lepidoptera) bewaard als bevroren gedragsinsluiting in Baltische Amber (Eoceen)
- ABSTRACT
- INTRODUCTION
- MATERIALEN EN METHODEN
- SYSTEMATISCHE PALAEONTOLOGIE
- DISCUSSIE
- Random voorkomen of een bewaard gebleven paring?
- Taphonomie
- Seksueel dimorfisme
- Habitat
- CONCLUSIE
- ACKNOWLEDGMENTS
Parende nachtvlinders (Tineidae, Ditrysia, Lepidoptera) bewaard als bevroren gedragsinsluiting in Baltische Amber (Eoceen)
Thilo C. Fischer en Marie K. Hörnig
Artikelnummer: 22.1.7
https://doi.org/10.26879/829
Copyright Paleontological Society, February 2019
Biografieën van auteurs
Plain-language and multi-lingual abstracts
PDF version
Inzending: 22 oktobere 2017. Acceptatie: 23 januari 2019
{flike id=2421}
ABSTRACT
Het onderzoek van barnsteeninsluitsels biedt een uniek inzicht in de biologie van dieren die miljoenen jaren geleden leefden, vanwege hun uitzonderlijke conservering in bijna levensechte omstandigheden. Dit geldt niet alleen voor morfologische aspecten, ook gedragsaspecten kunnen uit deze fossielen worden afgeleid.
Hier worden Microlepidoptera gemeld uit Eoceen Baltisch barnsteen, hoogstwaarschijnlijk als een eerste voorkomen van fossiele Lepidoptera gevonden in copula. De motten behoren tot de Tineidae (de kleermotten) en worden beschreven als een nieuwe soort en een nieuw genus, Forcepsites michalskii. Het klemorgaan van het mannetje is vrij ongewoon voor motten uit Baltisch barnsteen, met een ventraal en dorsaal deel, elk eindigend in twee parallelle, doornachtige uitsteeksels, en schijnbaar niet gevormd door de valvae. Het naast elkaar voorkomen van mannetjes en vrouwtjes, waarschijnlijk in copula, maakte het mogelijk hun sexueel dimorfisme rechtstreeks te bestuderen, en er zijn aanwijzingen dat de habitat van deze soort de plaats is waar hars wordt geproduceerd in het Baltische barnsteenwoud. Uit het sexueel dimorfisme van de antennes en de oogmorfologie wordt afgeleid dat bij deze soort het vrouwtje het roepende geslacht was voor de paring.
Thilo C. Fischer. Förderverein der Bayerischen Staatssammlung für Paläontologie und Historische Geologie München Richard-Wagner-Straße 10, D-80333 München, Duitsland.
Marie K. Hörnig. Universiteit Greifswald, Zoölogisch Instituut en Museum, Cytologie en Evolutionaire Biologie, Soldmannstr. 23, D-17489 Greifswald, Duitsland.
KEYWORDS: Kledingmotten; habitat in copula; Microlepidoptera; nieuwe soorten en genus; sexueel dimorfisme
Eindcitaat: Fischer, Thilo C, en Hörnig, Marie K. 2019. Parende nachtvlinders (Tineidae, Ditrysia, Lepidoptera) bewaard als bevroren gedragsinsluiting in Baltische Amber (Eoceen). Palaeontologia Electronica 22.1.7A 1-11. https://doi.org/10.26879/829
palaeo-electronica.org/content/2019/2421-mating-moths
Copyright: Februari 2019 Paleontological Society.
Dit is een open access artikel verspreid onder de voorwaarden van Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), die gebruikers toestaat om het materiaal te kopiëren en opnieuw te distribueren in elk medium of formaat, op voorwaarde dat het niet wordt gebruikt voor commerciële doeleinden en de oorspronkelijke auteur en bron worden gecrediteerd, met indicaties als er wijzigingen zijn aangebracht.
creativecommons.org/licenses/by-nc-sa/4.0/
http://zoobank.org/BCF64D89-6879-4805-8021-7CE926CE4B12
INTRODUCTION
Parende insecten worden zelden in barnsteen aangetroffen als gevallen van het zogenaamde ‘bevroren gedrag’ (Arillo, 2007, en verwijzingen daarin). Dergelijke zijn gerapporteerd uit Eoceen Baltisch barnsteen, bijvoorbeeld uit de groepen Diptera, Coleoptera, Hemiptera, Hymenoptera en vrij zelden Trichoptera (Weitschat en Wichard, 2002; Weitschat, 2009; Boucot en Poinar Jr., 2010; Gröhn, 2016).
In Baltisch barnsteen zijn Lepidoptera over het algemeen zeldzaam en komen voor bij ongeveer één op 250 inclusies (ter vergelijking: Dipteranen vertegenwoordigen ongeveer 64% van alle insecten in Baltisch barnsteen, Lepidoptera ongeveer 0,4 %; Hoffeins en Hoffeins 2003; Gröhn 2016). Net als bij andere insecten zijn in copula bewaarde imago’s veel zeldzamer en Lepidoptera in copula zijn nog niet beschreven, ook niet in andere ambers of uit niet-amber insectenlocaties (Sohn et al., 2012; Sohn en Lamas, 2013). Een in copula voorkomen maakt bonafide verwantschap van beide geslachtsvormen met elkaar en beide samen tot een aparte soort mogelijk, en daarmee de precieze beschrijving van seksueel dimorfisme van een soort. Anderzijds kan seksueel dimorfisme van fossiele insecten alleen met statistische middelen worden bestudeerd wanneer vondsten van een soort overvloedig zijn, zoals bij veel Nematocera (bijv, Borkent, 1995).
Tineidae zijn zeer divergente basale vertegenwoordigers van de lepidoptera groep Ditrysia (Regier et al., 2014), hun larven voeden zich met een breed scala van substraten (detritus, schimmels, korstmossen, of keratine) (Scoble, 1995), vandaar dat ze generalist zijn (85,7%) (Menken et al., 2009). Als insluitsels in Baltisch barnsteen vertegenwoordigen Tineidae ongeveer een kwart van alle vondsten van lepidoptera (Skalski, 1977). Hier wordt een zeldzame en nieuwe soort Tineidae beschreven met zijn geslachtsvormen van een vermoedelijke in copula voorkomen.
MATERIALEN EN METHODEN
Het barnsteenspecimen werd gevonden door en gekocht van Artur Michalski (Wroclaw, Polen). Het werd geconserveerd door behandeling met een acrylvernis en is afkomstig uit de collectie van de auteur (TF) met collectienummer 7131. Het werd bewaard bij constante temperatuur in plastic plastic zakken in metalen dozen, met uitsluiting van zuurstof en licht. Het zal worden gedeponeerd in de Bayerische Paläontologische Staatssammlung München met het nummer SNSB-BSPG 2017 I 113.
Het specimen werd gefotografeerd met een Canon EOS 70d uitgerust met een Canon MP-E 65 mm objectief en een Canon Twin Flash MT-24. Om reflecties te vermijden, werden polarisatiefilters gemonteerd en afgesteld voor de flitsers en het camera-objectief (kruisgepolariseerd licht). Om de convexiteit en concaviteiten van het amberkleurige oppervlak te verminderen, werd een druppel glycerine aangebracht en afgedekt met een dekglaasje. De opgenomen beeld z-stacks werden samengesmolten met Combine ZP en verder verwerkt met Adobe Photoshop CS 3. De insluitsels konden niet worden bestudeerd met microcomputer-tomografie, vanwege de aanwezigheid van metaalachtige, schijnbaar pyritische insluitsels, die sterke artefacten veroorzaken.
SYSTEMATISCHE PALAEONTOLOGIE
Systematiek (volgens van Nieukerken et al., 2011)
Orde LEPIDOPTERA Linnaeus, 1758
Clade DITRYSIA Börner, 1925
Superfamilie TINEOIDEA Latreille, 1810
Familie TINEIDAE Latreille, 1810
Genus Forcepsites nov. gen.
zoobank.org/DDB6884D-845A-4A87-981E-A2953ADD8964
Etymology. “Forceps”, latijn voor tang, verwijzend naar het mannelijke grijporgaan.
Soort Forcepsites michalskii nov. spec.
zoobank.org/5494D58E-7773-473C-AFB2-140DD049226
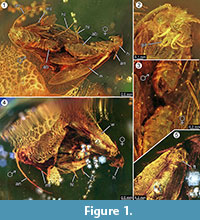 Holotypus. Exemplaar ex coll. Fischer no. 7131, Figure 1.2
Holotypus. Exemplaar ex coll. Fischer no. 7131, Figure 1.2
Locus typicus. Barnsteenmijn van Yantarni, GUS
Stratum typicum. “Blaue Erde” (bovenste Eoceen – onderste Oligoceen)
Etymologie. De soortnaam michalskii is gekozen als dank aan Artur Michalski voor het leveren van dit specimen.
Repository. Bayerische Staatssammlung für Paläontologie und Geologie, München, Duitsland; toetredingsnummer SNSB-BSPG 2017 I 113.
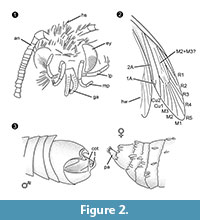
Diagnose van het genus. Monotypisch geslacht met als bepalend kenmerk het omklemorgaan van het mannetje (figuur 2.3) met een ventraal en dorsaal deel, elk eindigend in twee parallelle, doornvormige uitsteeksels.
 Diagnose van de soort. De respectieve kwalitatieve en kwantitatieve kenmerken van de vrouwelijke en mannelijke imago’s (figuur 1) werden bepaald en zijn samengevat in tabel 1.
Diagnose van de soort. De respectieve kwalitatieve en kwantitatieve kenmerken van de vrouwelijke en mannelijke imago’s (figuur 1) werden bepaald en zijn samengevat in tabel 1.
Differentiële diagnose. De combinatie van tekens kleine proboscis met gescheiden galeae, lange en gevouwen maxillaire palpen met vijf segmenten, naar boven gerichte en licht gebogen labiale palpen met laterale borstelharen, kop met veel rechtopstaande haarachtige schubben suggereren sterk een verwantschap met Tineidae, maar geen van deze is apomorf, evenmin is een dergelijke apomorfie momenteel bekend voor volwassen Tineidae (Regier et al., 2014). De nerven R4 en R5 zijn niet vergroeid / gesteeld, wat indicatief zou zijn voor Gelechioidea (Gelechiidae en Oecophoridae) (Scoble, 1995), de andere grote groep Lepidoptera die in Baltisch barnsteen is aangetroffen. De soort lijkt op geen enkele andere beschreven vertegenwoordiger van Tineidae of andere families uit Baltisch barnsteen (Rebel, 1934, 1935; Kuznezov, 1941; Skalski, 1973; Kozlov, 1987, 1988; Sohn et al., 2012; Sohn and Lamas, 2013), vooral wat betreft het mannelijke genitale apparaat. Het meest gelijkend lijkt Tineosemopsis decurtatus Skalski 1974 (vrouwtje bekend), maar verschilt in de afwezigheid van rechtopstaande haren aan de vertex en zijn kleinere ogen. Tillyardinea eocaenica Kusnezov 1941 (vrouwtje), die rechtopstaande schubben op de vertex heeft en een gelijkaardige venatie, verschilt in zijn frons en labiale palpen. Simulotinea intermedia Skalski 1977 heeft een gelijkaardige habitus maar verschilt ook door de aanwezigheid van een tweesplitsende ader, die de grote cel in voorvleugel (M2+3) verdeelt.
Taphonomie. Mannetje en vrouwtje van Forcepsites michalskii zijn nabij elkaar gefossiliseerd en met het achterste deel van het achterlijf naar elkaar toe gericht. De twee insluitsels bevinden zich in een cilindrisch stuk niet-geautoclaveerd Baltisch barnsteen met een diameter van 15 mm en een hoogte van 11 mm dat op ongeveer de helft van zijn hoogte is verdeeld door een ongelijke rand tussen twee lagen harsvloei. Deze rand vertoont een laag pyriet als deel van zijn vlak. Eén harsvloei bevat de twee motten die hun respectieve dorsale zijden aan deze rand blootstellen. Van de rand van de vleugel van het vrouwtje zijn lange schubben bewaard gebleven. De vrouwelijke imago heeft twee poten verloren, en haar borststuk is beschadigd. De andere stroom bevat niet-lineaire reeksen van belletjes. Beide individuen lijken pas uitgekomen te zijn, te oordelen naar het volledige kleed van schubben op de vleugels en het feit dat de vleugels niet versleten zijn. Er konden geen reukorganen worden vastgesteld op de vleugels, de buik of de poten. De enige syninclusies die bij de Microlepidoptera bewaard zijn gebleven, zijn trichomen, die typisch zijn voor Baltisch barnsteen.
DISCUSSIE
Random voorkomen of een bewaard gebleven paring?
De ingesloten motten zijn mannelijke en vrouwelijke vertegenwoordigers van dezelfde soort en staan dicht bij elkaar met de buikspieren naar elkaar toe gericht. Toch zijn de geslachtsdelen van de motten niet in direct contact, maar in een nauwe positie. De dorsale/ventrale oriëntatie van het mannetje en het vrouwtje verschilt enigszins, wat veroorzaakt wordt door een lichte rotatie binnen de lichaamsas (vooral bij het mannetje). De oriëntatie van het achterste deel van het achterlijf van beide motten is naar elkaar toe.
Er zijn verschillende gevallen bekend, waarbij de situatie van opsluiting in de nog vloeibare hars een stressrespons kan opwekken, zoals ovipositie (Weitschat, 2009). De inductie van paringsgedrag door de harsstroom is in het algemeen niet aannemelijk, maar de situatie van opsluiting toont aan dat lichte bewegingen binnen de verse hars mogelijk waren.
Er is ook de vraag of deze bevinding ook een toevallige coïncidentie zou kunnen vertegenwoordigen. De kans op zo’n toevallige coëxistentie kan ruw geschat worden. Onder de insecteninsluitsels komen motten voor met een frequentie van ongeveer één op 250 insluitsels in Baltisch barnsteen (Krzeminskaet al., 1991 (0,6%); Hoffeins en Hoffeins, 2003 (0,4%); Sontag, 2003 (0,27%), en eigen waarneming). Voor een individuele barnsteen waarin een mot voorkomt, is de kans op een toevallig samen voorkomen van een andere mot eenvoudigweg 1:250 (motten komen niet in zwermen voor). Het samen voorkomen van hetzelfde taxon, of een overvloedig taxon dat verwisseld zou kunnen worden, wordt geschat op 1:10 in het beste geval (Skalski, 1977), en nog eens ongeveer 1:2 voor het hebben van verschillende geslachten. Rekening houdend met de verdere kansen op nauwe groepering, zowel uitlijning van de lichaamsassen, als een geschikte lichaamsoriëntatie, is de kans op een toevallige coïntroductie die verkeerd geïnterpreteerd zou kunnen worden als een paring, veel kleiner dan de waargenomen frequentie als één op 920 lepidoptera in de collectie van de bestudeerde auteur.
Concluderend is de meest waarschijnlijke interpretatie voor deze bevinding dat de motten in copula waren, en de genitaliën in contact waren geweest, maar dat de individuen enigszins van elkaar waren afgeleid als gevolg van de harsstroom (zie discussie hieronder).
Taphonomie
De relevantie van een zorgvuldige analyse van harsstromen voor de taphonomie van barnsteeninsluitingen is aangetoond door Coty et al. (2014) met hun studie van een mier – termiet interactie bewaard gebleven in Mexicaans Chiapas barnsteen.
Het optreden van een in copula gebeurtenis wordt sterk ondersteund door het vinden van lange schubben afgeleid van de rand van de vleugel van het vrouwtje, die over enige afstand in de richting van het mannetje afdreven. Dit zou kunnen wijzen op een sterke laminaire harsstroom op deze plaats, die langs hun gemeenschappelijke as bewoog, en hen voor de nu waargenomen korte afstand uit elkaar getrokken zou kunnen hebben. Tijdens de paring van nachtvlinders kan het mannetje het contact met de paringsplaats verliezen en alleen door het omklemmende orgaan aan het vrouwtje worden vastgemaakt, waarbij alleen het vrouwtje contact maakt met het oppervlak. Dit gedrag en de weerstand van de plotselinge harsstroom zouden de reden kunnen zijn waarom het vrouwtje haar beide achterpoten heeft verloren. Aan de rugzijde van de motten, die aan het convexe oppervlak van deze eerste laminaire harsstroom was blootgesteld, zijn de vleugel van het vrouwtje en de onderliggende thorax gedeeltelijk beschadigd (figuur 1.5). Vermoedelijk zijn deze beschadigde plekken voedingssporen veroorzaakt door andere insecten, waarschijnlijk mieren zoals vaak wordt waargenomen in barnstenen specimens. Na de eerste harsstroom als inbeddingsgebeurtenis zullen de dorsale zijden dus enige tijd bloot hebben gelegen, lang genoeg om predatoren in staat te stellen over dit vaste oppervlak te lopen. De tweede harsstroom, die deze zijde later bedekte, zou een snelle en turbulente harsstroom geweest kunnen zijn, zoals blijkt uit een reeks kleine belletjes in onregelmatige krommingen die daarin gevonden zijn. Dit zou een typisch taphonomisch proces zijn voor een zogenaamde “Schlaube” van Baltisch barnsteen (Gröhn, 2016).
Seksueel dimorfisme
Seksueel dimorfisme is moeilijk waar te nemen bij fossiele soorten, waarvan meestal slechts enkele exemplaren bekend zijn. Kenmerken die niet beïnvloed worden door het geslacht moeten diagnostisch zijn om een zekere soortbepaling mogelijk te maken als basis voor de verdere differentiatie tussen de geslachten en de opheldering van seksueel dimorfisme. Bij sterk seksueel dimorf levende soorten, b.v. met brachypteres zoals bij veel Psychidae (Scoble, 1995), is het nauwelijks mogelijk om beide geslachten tot één soort te rekenen. Vandaar dat alleen gevallen van zwak sexueel dimorfisme betrouwbaar kunnen worden herkend in fossiele soorten met behulp van statistische middelen en zonder ze te vinden in copula. Dit onderstreept het belang van geïdentificeerd paringsgedrag bij fossiele soorten voor de beschrijving van zulke gevallen van sterk sexueel dimorfisme. Dergelijke bewaard gebleven vondsten van fossiele paringen kunnen ook de correlatie goedkeuren van eerder bekende sexueel dimorfe vormen om tot één soort te behoren, zelfs zonder de mogelijkheid van veel statistische ondersteuning. Er is enige kans dat een bewaard gebleven paring een interspecifieke of intergenerieke paring vertegenwoordigt, zoals deze bekend zijn van nachtvlinders en vlinders (Ganyard en Brady, 1972; Deering en Scriber, 2002). Deze verschijnselen komen echter veel minder vaak voor dan intraspecifieke paring en het is nauwelijks waarschijnlijk dat ze als een zeldzame fossiele insluiting zullen worden gevonden.
Een niet-allometrisch sexueel dimorfisme dat is waargenomen bij Forcepsites michalskii is de gescheiden galeae in de proboscis bij het mannetje. Dit kan duiden op een niet-functionele toestand, waarbij de mannelijke imago’s van deze soort niet-voedend zijn, zoals gezien wordt bij veel nog levende (mannelijke) Psychidae met een rudigiale of afwezige proboscis (Scoble, 1995). Allometrische sexuele dimorfismen die hier gevonden worden zijn: (1) Het vrouwtje is groter dan het mannetje, zowel in lichaamsgrootte als in vleugellengte. De respectieve verhoudingen zijn echter vrijwel gelijk. (2) Bij het vrouwtje is het achterlijf relatief en absoluut breder in vergelijking met het mannetje. (3) De antennes van het mannetje zijn veel groter en dikker dan bij de vrouwelijke imago. Bij verschillende nog levende soorten zijn de antennen seksueel dimorf (Scoble, 1995), de goed ontwikkelde mannelijke antennen dienen voor de chemosensatie van vrouwelijke feromonen. (4) De maxillaire palpen van het mannetje zijn tweemaal zo dik als bij het vrouwtje en hebben een geribbelde structuur. (5) De ogen van het mannetje zijn ook groter, maar het aantal ommatidia is ongeveer gelijk. Grotere ogen zullen alleen meer licht ontvangen en zouden gevoeliger kunnen zijn; de resolutie zou echter hetzelfde zijn.
Herkenning op lange afstand van roepende vrouwelijke nachtvlinders vindt plaats via mannelijke chemoreceptoren terwijl herkenning op korte afstand in plaats daarvan berust op visuele identificatie (Groot, 2014). Bij Forcepsites michalskii lijkt het er ook op dat het vrouwtje het roepende geslacht was -zoals gesuggereerd wordt door het grotere antenneapparaat en de ogen van het mannetje.
Habitat
Op basis van de hierboven gepresenteerde discussie is het duidelijk dat in copula gevangen individuen alleen kunnen voorkomen op de plaats waar deze soort leefde en paarde. Afzonderlijke individuen kunnen door de wind worden weggedreven of niet-habitat plaatsen bezoeken, zoals aquatische insecten die zelden in Baltisch barnsteen worden aangetroffen (Wichardet al., 2009); voor parende individuen daarentegen zou dit nauwelijks het geval kunnen zijn zonder gescheiden te worden. Er lijkt in de literatuur geen bewijs te zijn voor paring van nachtvlinders tijdens de vlucht. Bij de meeste Lepidoptera en bij overwegend nachtactieve microlepidoptera (nachtvlinders) daarentegen “roepen” de vrouwtjes de mannetjes op met feromonen en bepalen zo de paringsplaatsen (Groot, 2014). Het kan zijn dat ze vliegen voor de paring, maar in de meeste gevallen lijken ze sessiel te zijn en uit te waaieren voor de feromoonverspreiding. De seksen die roepen of zoeken kunnen bij sommige taxa echter ook omgekeerd zijn, maar voor Tineidae wordt gemeld dat het vrouwtje het roepende geslacht is (Wagner en Rosovsky, 1991; Scoble, 1995). In elk geval was de plaats van de harsstroom, d.w.z. de biotoop met de barnsteen producerende bomen, hoogstwaarschijnlijk ook de habitat van de hier beschreven kleermot.
CONCLUSIE
Een tineoïde mot met een opmerkelijk mannelijk genitaal apparaat wordt beschreven als de nieuwe soort en genus Forcepsites michalskii uit Eoceen Baltisch barnsteen. Vrouwtje en mannetje zijn samen bewaard in nauwe positie en met het achterlijf naar elkaar toe gericht, wat suggereert dat deze motten tijdens de paring in de verse hars gevangen zaten. Een sterke laminaire harsstroom, die wordt aangegeven door verschillende afgedreven delen van het lichaam van het vrouwtje, zou hen over de korte afstand die nu is waargenomen, uit elkaar hebben kunnen trekken. Dit specimen zou dus het eerste voorbeeld kunnen zijn van fossiele Lepidoptera gevangen in copula.
Aannemelijk is dat Forcepsites michalskii een bosbewonende soort was, aangezien individuen gevangen in copula alleen gedocumenteerd kunnen worden van de plaats waar deze soort leefde en paarde.
Vrouwtjes en mannetjes van F. michalskii vertonen sexuele dimorfismen in lichaamsgrootte, vleugellengte, verhouding van het achterlijf, de grootte van de samengestelde ogen, en de morfologie van de maxillaire palpen, evenals van de antennes. Gebaseerd op de duidelijk grotere en dikkere antennes van het mannetje, alsmede op zijn grotere ogen, wordt gesuggereerd dat het vrouwtje het roepende geslacht zou kunnen zijn bij deze soort, zoals ook gerapporteerd bij bestaande Tineidae soorten.
ACKNOWLEDGMENTS
A. Michalski (Wroclaw, Polen) wordt bedankt voor het leveren van dit uitzonderlijke specimen en het leiden van onze aandacht op het. M. Schwarz van het Oberösterreichisches Landesmuseum (Linz, Oostenrijk) was zo vriendelijk de screening van hun Lepidoptera-collectie te ondersteunen. MKH werd vriendelijk gefinancierd door een beurs van de Studienstiftung des deutschen Volkes en dankt S. Harzsch (Greifswald, Duitsland) voor zijn steun. Wij erkennen het waardevolle wetenschappelijke commentaar van M. Heikkilä en C. Haug (München, Duitsland) op het manuscript, evenals het commentaar van J. Krieger (Greifswald, Duitsland) en de steun van C. MacGraphery (Freising, Duitsland) en Y. Goldman (Collinsville, CT, VS) om de Engelse tekst te controleren als moedertaalsprekers. Verder danken wij twee anonieme reviewers voor hun nuttige commentaar.
Arillo, A. 2007. Paleoethologie: gefossiliseerde gedragingen in barnsteen. Geologica Acta, 5(2):159-166. https://doi.org/10.1344/105.000000301
Börner, C. 1925. Lepidoptera, Schmetterlinge, p. 358-387. In Brohmer, P. (ed.). Fauna von Deutschland. Ein Bestimmungsbuch unserer heimischen Tierwelt. Quelle und Meyer, Leipzig.
Borkent, A. 1995. Bijtende Muggen in het Krijt van Noord-Amerika (Diptera: Ceratopogonidae). Uitgeverij Backhuys, Leiden, Nederland.
Boucot, A.J. and Poinar Jr, G.O. 2010. Fossil Behavior Compendium. CRC Press, Boca Raton, p. 147-151.
Coty, D., Aria, C., Garrouste, R., Wils, P., Legendre, F. and Nel, A. 2014. The first ant-termite syninclusion in amber with CT-scan analysis of taphonomy. PLOS one, 9(8):e104410. https://doi.org/10.1371/journal.pone.0104410
Deering, M.D. and Scriber, J.M. 2002. Field bioassays show heterospecific mating preference asymmetry between hybridizing North American Papilio butterfly species (Lepidoptera: Papilionidae). Tijdschrift voor ethologie 20:25-33. https://doi.org/10.1007/s10164-002-0050-2
Fletcher, T.B. 1940. New generic names for microlepidoptera. Entomologist’s Record, 52:17-19.
Ganyard, M.C. and Brady, Jr.U.E. 1972. Interspecific attraction in Lepidoptera in the field. Annals of the Entomological Society of America, 65 (6): 1279-1282. https://doi.org/10.1093/aesa/65.6.1279
Gröhn, C. 2016. Insluitsels in Baltische amber. Wachholtz Verlag – Murmann Publishers, Kiel, Hamburg.
Groot, A.T. 2014. Circadiane ritmen van sexuele activiteiten bij motten: Een overzicht. Frontiers in Ecology and Evolution 2:43. https://doi.org/10.3389/fevo.2014.00043
Hoffeins, C. en Hoffeins, H.W. 2003. Studies over de abundantie van insluitsels in Baltisch en Bitterfeld barnsteen (Tertiair, Eoceen) uit niet-geselecteerde collecties, met speciale aandacht voor de orde Diptera. Studia Dipterologica, 10(2):381-392.
Kozlov, M.V. 1987. Nieuwe motachtige Lepidoptera uit Baltisch barnsteen. Paleontologisch Tijdschrift, 4:59-67.
Kozlov, M.V. 1988. Paleontologie van lepidoptera en problemen met de fylogenie van de orde Papilionida, p. 16-69. In Ponomarenko A.G. (ed.), The Mesozoic-Cenozoic Crisis in the Evolution of Insects. Academie van Wetenschappen, Moskou.
Krzeminska, E., Krzeminski, W., Haenni, J.-P., and Dufour, C. 1991. Les Fantomes De L’Ambre. Muséed’histoire naturelle de Neuchâtel, Neuchâtel, Zwitserland.
Kusnezov, N. 1941. Een revisie van Amber Lepidoptera. Paleontologisch Instituut, USSR Academie van Wetenschappen, Moskou & Leningrad.
Latreille, P.A. 1810. Algemene beschouwingen over de natuurlijke orde der dieren, die de klassen der schaaldieren, spinachtigen en insecten uitmaken: met een methodische tabel van hun geslachten, gerangschikt in families. Parijs, Schoell.
Menken, S.B.J., Boomsma, J.J. en van Nieukerken, E.J. 2009. Grootschalige evolutionaire patronen van waardplantassociaties bij de Lepidoptera. Evolution, 64(4):1098-1119. https://doi.org/10.1111/j.1558-5646.2009.00889.x
Rebel, H. 1934. Bernstein-Lepidopteren (aus der Sammlung Bachofen-Echt). Palaeobiologica, 6:1-16.
Rebel, H. 1935. Bernstein-Lepidopteren. Entomologische Zeitschrift Iris, 49:162-186.
Regier, J.C., Mitter, C., Davis, D.R., Harrison, T.L., Sohn J.-C., Cummings, M.P., Zwick, A., and Mitter, K.T. 2014. A molecular phylogeny and revised classification for the oldest Ditrysian moth lineages (Lepidoptera: Tineoidea), with implications for ancestral feeding habits of the mega-diverse Ditrysia. Systematische entomologie, 40(2). https://doi.org/10.1111/syen.12110
Scoble, M.J. 1995. The Lepidoptera – Form, Function and Diversity. The Natural History Museum / Oxford University Press, Oxford.
Skalski, A.W. 1973. Studies over de Lepidoptera uit fossiele harsen. Deel II. Epiborkhausenites obscurotrimaculatus gen. et sp. nov. (Oecophoridae) en een Tineid-mot ontdekt in Baltisch barnsteen. Acta Palaeontologica Polonica, 18(1):153-160.
Skalski, A.W. 1974. Zwei neue Gattungen und Arten der Familie Tineidae aus dem Baltischen Bernstein. Studien an Lepidopteren aus fossilen Harzen V. Beiträge zur Entomologie, 1/4(24):97-104.
Skalski, A.W. 1977. Studies over de Lepidoptera uit fossiele harsen. Deel I. Algemene opmerkingen en beschrijvingen van nieuwe geslachten en soorten van de families Tineidae en Oecophoridae uit Baltisch barnsteen. Prace Museum Ziemi, 26:3-24.
Sohn, J.-C., Labandeira, C., Davis, D., and Mitter, C. 2012. An annotated catalog of fossil and subfossil Lepidoptera (Insecta: Holometabola) of the world. Zootaxa, 3286:1-132.
Sohn, J.-C. and Lamas, G. 2013. Corrections, additions, and nomenclatural notes to the recently published world catalog of fossil and subfossil Lepidoptera. Zootaxa, 3599:395-399.
Sontag, E. 2003. Animal inclusions in a sample of unselected Baltic amber. Acta Zoologica Cracoviensia, 46 (suppl.-Fossiele Insecten):431-440.
van Nieukerken, E.J., Kaila, L., Kitching, I.J., Kristensen, N.P., Lees, D.C., Minet, J., Mitter, C., Mutanen, M., Regier, J.C., Simonsen, T.J., Wahlberg, N., Yen, S.-H., Zahiri, R., Adamski, D., Baixeras, J., Bartsch, D., Bengtsson, B.Å., Brown, J.W., Bucheli, S.R., Davis, D.R., De Prins, J., De Prins, W., Epstein, M.E., Gentili-Poole, P., Gielis, C., Hätteschwiler, P., Hausmann, A., Holloway, J.D., Kallies, A., Karsholt, O., Kawahara, A.Y., Koster, S.J.C., Kozlov, M.V., Lafontaine, J.D., Lamas, G., Landry, J.-F., Lee, S., Nuss, M., Park, K.-T., Penz, C., Rota, J., Schintlmeister, A., Schmidt, B.C., Sohn, J.-C, Solis, M.A., Tarmann, G.M., Warren, A.D., Weller, S., Yakovlev, R.V., Zolotuhin, V.V., and Zwick, A. 2011. Orde Lepidoptera Linnaeus, 1758. In Zhang, Z.-Q. (ed.), Dierlijke biodiversiteit: An outline of higher-level classification and survey of taxonomic richness. Zootaxa, 3148:212-221.
von Linné, C. 1758. vertaald door William Turton (1806). Deel 1. Een algemeen systeem van de natuur: Through the Three Grand Kingdoms of Animals, Vegetables, and Minerals, Systematically Divided into their Several Classes, Orders, Genera, Species, and Varieties. Lackington, Allen, and Co, London.
Wagner, D.L. and Rosovsky, J. 1991. Mating systems in primitive Lepidoptera, with emphasis on the reproductive behavior of Korscheltellus gracilis (Hepialidae). Zoological Journal of the Linnean Society, 102:277-303. https://doi.org/10.1111/j.1096-3642.1991.tb00003.x
Weitschat, W. and Wichard, W. 2002. Atlas van planten en dieren in Baltisch barnsteen. Verlag Friedrich Pfeil, München.
Weitschat, W. 2009. Jäger, Gejagte, Parasiten und blinde Passagiere – Momentaufnahmen aus dem Bernsteinwald. Denisia, 26(86):243-256.
Wichard, W., Gröhn, C., and Seredszus, F. 2009. Aquatic Insects in Baltic Amber. Verlag Kessel, Remagen-Oberwinter.