Cleavage se referă la modelul stereotipizat al diviziunilor mitotice timpurii care împarte volumul mare de citoplasmă a oului. Zigotul timpuriu este unic prin faptul că este atât de mare. Cele mai multe celule trec printr-o perioadă de creștere între ciclurile de mitoză, dar acest lucru nu este valabil pentru blastomerele în stadiul de clivaj timpuriu. Cu fiecare diviziune, celulele devin mai mici. Acest model rapid de diviziune celulară fără creștere concomitentă se oprește brusc în stadiul numit tranziția mijlocie a blastomului, în care nucleul zigotului preia controlul ciclului celular.
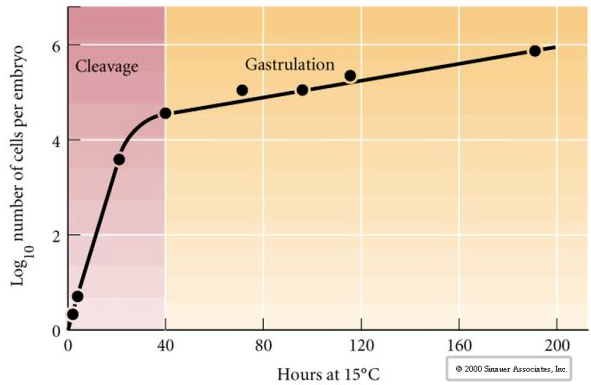 Există unele dovezi că un factor matern, poate un regulator transcripțional, este responsabil pentru acest model rapid timpuriu de diviziuni de clivaj. Prin modificarea artificială a raportului dintre ADN citoplasmatic și ADN nuclear se poate modifica momentul tranziției de la jumătatea blastulei. Tranziția midblastula se referă la momentul în care are loc trecerea majoră de la expresia genelor materne la cele zigotrice.
Există unele dovezi că un factor matern, poate un regulator transcripțional, este responsabil pentru acest model rapid timpuriu de diviziuni de clivaj. Prin modificarea artificială a raportului dintre ADN citoplasmatic și ADN nuclear se poate modifica momentul tranziției de la jumătatea blastulei. Tranziția midblastula se referă la momentul în care are loc trecerea majoră de la expresia genelor materne la cele zigotrice.
Fertilizarea la unele specii duce la mișcări citoplasmatice radicale care sunt esențiale pentru a se asigura că determinanții citoplasmatici sunt localizați în pozițiile corecte în raport cu evenimentele ulterioare de clivaj.
PATRONURILE CLASAMENTULUI EMBRYONIC
Patronajul clivajului embrionar este determinat atât de poziția fusurilor mitotice, cât și de cantitatea și distribuția de gălbenuș. Gălbenușul are tendința de a inhiba clivajul. Acesta o încetinește sau chiar împiedică clivarea completă. Gălbenușul este o adaptare a acelor animale care parcurg mai mult sau mai puțin timp embriogeneza izolate de orice sursă de hrană. Unele animale, cum ar fi ariciul de mare, au relativ puțin gălbenuș, deoarece se dezvoltă rapid într-o formă larvară care înoată liber și care capătă nutrienți din mediul înconjurător. Alte animale, cum ar fi marsupialele, se nasc prematur, dar primesc hrană într-o pungă parentală. Mamiferele placentare dezvoltă un organ specializat prin care embrionul este hrănit pe tot parcursul dezvoltării și astfel au, de asemenea, puțin gălbenuș.
Tipurile de ouă în funcție de caracteristicile gălbenușului sunt descrise astfel:
Isolecitale: gălbenuș rar distribuit uniform, de ex. ariciul de mare, șoarecele
Mesolecitale: cantitate moderată de gălbenuș, adesea distribuit neuniform, de ex, broasca
Telolecitală: gălbenuș dens concentrat la un capăt, de exemplu, pasăre, reptilă
Centrolecitală: gălbenuș concentrat în mijlocul oului, de exemplu, musca
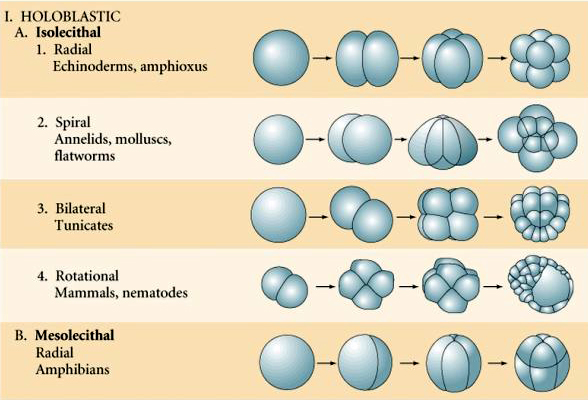
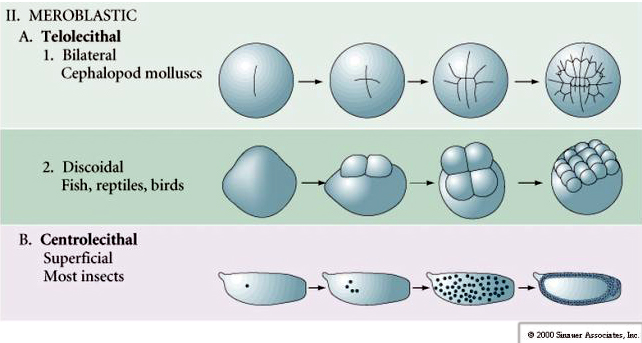
Multe ouă sunt polarizate cu un pol bogat în gălbenuș, numit pol vegetal și un pol sărac în gălbenuș numit pol animal, de exemplu, broasca. Nucleul zigotului este, în general, deplasat spre polul animal. Zigoții cu o cantitate relativ mică de gălbenuș (izolecitali și mezolecitali) se scindă HOLOBLASTICAL. Șanțul de clivaj se întinde pe tot parcursul oului. În timp ce zigoții telolecitali și centrolecitali sunt supuși clivajului MEROBLASTIC, unde planul de clivaj se extinde doar până la gălbenușul acumulat. La ouăle centrolecitale (multe ouă de insecte) clivajul este meroblastic și superficial, în timp ce la ouăle telolecitale (păsări și pești) clivajul este discoidal
Există mai multe tipuri de simetrie a clivajului observate în natură: radială (echinoderme, amfibieni), spiralată (moluște, anelide), bilaterală (ascidieni,tunicate), rotațională (mamifere). Cele două figuri de mai jos prezintă exemple de simetrii ale clivajului holoblastic și meroblastic.
CLIVAJ HOLOBLASTIC RADICAL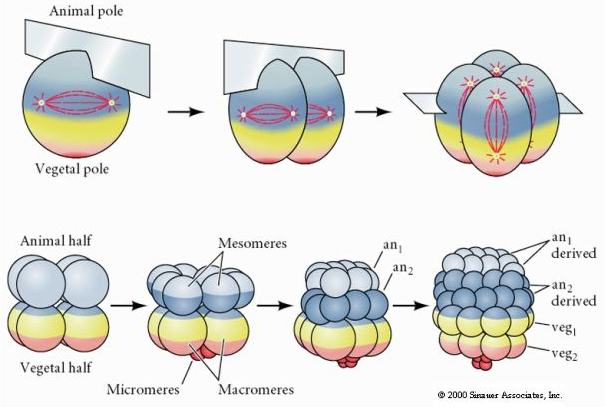
Excelent film al clivajului ariciului de mare din cartea lui Rachel Fink „A Dozen Eggs”.
Aricii de mare au, de asemenea, un clivaj holoblastic radial, dar cu unele diferențe interesante. Primul clivaj este meridional. al doilea clivaj este meridional. Al treilea clivaj este ecuatorial Al patrulea clivaj este meridional, dar în timp ce cele patru celule de la polul animal se împart în mod egal pentru a da naștere la opt blastomere animale de dimensiuni egale numite MESOMERE, celulele vegetale se împart asimetric de-a lungul planului ecuatorial pentru a da 4 MACROMERE mari și 4 MICROMERE mult mai mici la polul vegetal. La a cincea diviziune, MESOMERES se împart pe plan ecuatorial pentru a da două niveluri de opt MESOMERES an1 și an2, MACROMERES se împart pe plan meridional, formând un nivel de opt celule sub an2, iar MICROMERES se împart pentru a da un grup de celule sub stratul veg1. Cele șase diviziuni sunt toate ecuatoriale, dând un strat veg2. A șaptea diviziune este toată meridională, dând o blastulă cu 128 de celule.
Ce determină aceste modele de clivaj? Sunt ele dependente de clivajul anterior și se derulează ca o bandă sau sunt determinate de un ceas intrinsec? În 1939, Horstadius a inhibat unul sau două dintre primele trei clivaje și a constatat că apariția micromerelor a avut loc la momentul potrivit, indiferent de istoricul clivajelor
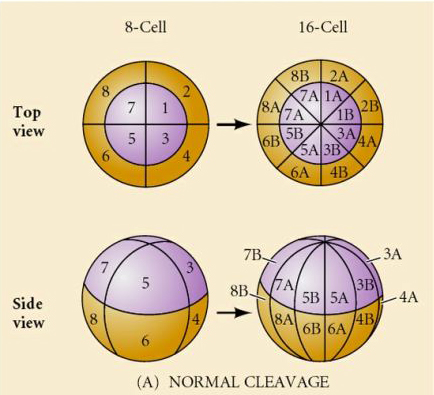
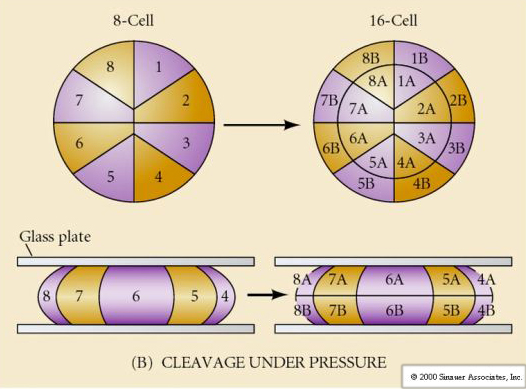
Concluzia acestor experimente este că există un factor în polul vegetal al oului care determină formarea micromerelor și, mai mult, că trebuie să existe un ceas molecular care începe la activarea oului. Ceasul este independent de evenimentul propriu-zis de clivaj.
Blastulele cu 128 de celule sunt o minge destul de liberă de celule care înconjoară un blastocel gol. Mingea are un strat celular gros de un singur strat celular, toate celulele fiind în contact cu stratul hialin extern și cu lichidul intern al blastococelului. În acest stadiu al dezvoltării, celulele încep să formeze joncțiunile strânse caracteristice unui epiteliu. Blastococelul central este acum izolat de mediul extern. Blastomerii continuă să se dividă cu axa lor paralelă cu stratul hialin, rămânând un epiteliu cu grosimea de o celulă. Blastococelul continuă să se mărească.
Două teorii încearcă să explice modelul de mărire a blastocistului
1. Teoria osmotică sugerează că ionii și proteinele sunt secretate în blastococel de către blastomeri și acest lucru duce la o creștere a presiunii datorită fluxului osmotic de apă. Această presiune ar fi apoi responsabilă de alinierea axei mitotice a blastomerilor și de mărirea blastococelului.
2. Teoria alternativă a lui Wolpert și a colegilor săi sugerează că, de fapt, interacțiunile adezive dintre blastomeri și dintre blastomeri și stratul hialin sunt cele care aliniază axele mitotice. Adică aderența la hialină este cea mai mare, urmează aderența la alți blastomeri și, în cele din urmă, interacțiunea cu peretele blastocoelului este cea mai mică. Adeziunea dominantă cu stratul hialin forțează expansiunea blastocistului și a blastococelului.
Celulelor din blastulă le cresc cili pe suprafața lor exterioară, secretă o enzimă de ecloziune (hialinaza) și devin înotătoare libere.
Clevajul amfibian
Clevajul la mulți amfibieni este holoblastic cu simetrie radială, însă volumul mare de gălbenuș (mezolecitul său) interferează cu clevajul. La polul animal primul clivaj se desfășoară cu aproximativ 1mm/min, în timp ce prin polul vegetal se desfășoară de 50-100 de ori mai lent (.02mm/min). În timp ce primul clivaj este încă incomplet în regiunea vegetală gălbuie a oului, începe să aibă loc cel de-al doilea clivaj meridional.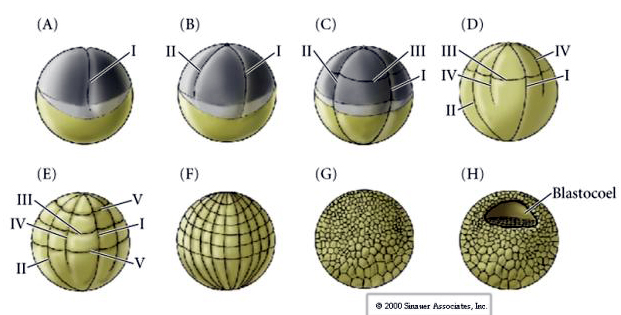
Cel de-al treilea clivaj este ecuatorial, dar, deoarece nucleele și astrele sunt deplasate spre animal, planul de clivaj, deși perpendicular pe axa vegetală animală, este de asemenea deplasat spre polul animal și nu împarte în mod egal blastomerii. Rezultă patru blastomere animale mai mici (denumite MICROMERE) și patru blastomere mari de la polul vegetal (denumite MACROMERE). Această scindare holoblastică inegală dă naștere unui pol animal cu diviziune mai rapidă, alcătuit din micromeri mai mici, și unui pol vegetal cu diviziune mai lentă, alcătuit din macromeri. În scurt timp, polul animal este compus din multe micromere mici, iar polul vegetal din câteva macromere mari, pline de gălbenuș. Deși formarea blastocelului începe odată cu primul clivaj, aceasta nu devine evidentă până în stadiul de 128 de celule.
Ce funcție îndeplinește blastocelul?
Blastocelul separă spațial celulele astfel încât acestea să nu se atingă între ele. Celulele de la acoperișul blastococelului devin în mod normal ectoderm. Dacă transplantați celulele din acoperișul blastococelului lângă celulele gălbui de la baza blastococelului, acestea se vor diferenția ca mezoderm. Derivații mezodermici sunt produși în mod normal din celulele adiacente precursorilor endodermici. O posibilitate pe care o vom explora în detaliu este aceea că celulele vegetale induc, prin interacțiuni celulă-celulă, celulele adiacente să devină mezoderme. Astfel, formarea blastocoelului poate fi necesară pentru a preveni interacțiunile „inductive” inadecvate între celulele timpurii ale blastocistului. A doua necesitate evidentă a blastococelului poate fi în timpul etapei ulterioare de dezvoltare, GASTRULAREA, în care celulele migrează în interiorul blastococelului.
CLASTOCelul de mamifere
Oulul de mamifer este eliberat din ovar în oviduct, unde este fecundat. Primul clivaj începe la aproximativ o zi după fertilizare în interiorul oviductului. În contrast puternic cu majoritatea animalelor, clivajul la mamifere poate fi foarte lent –1/zi.
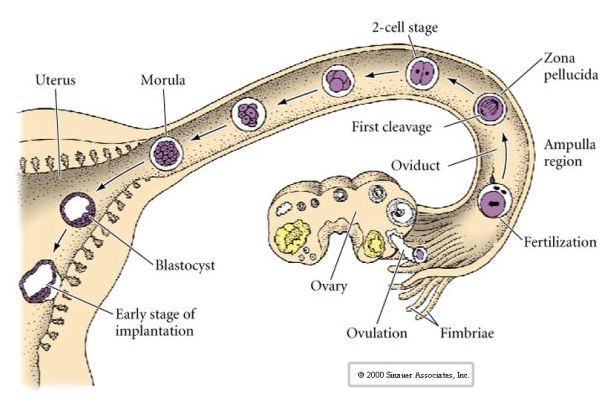
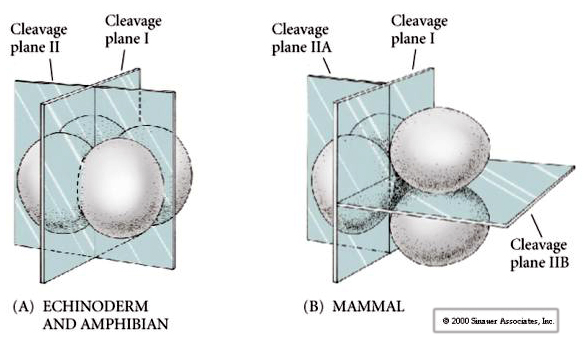 În plus, planurile de clivaj sunt oarecum diferite de cele ale altor animale. Primul clivaj este meridional, la fel ca în cazul ariciului de mare și al broaștei. Cu toate acestea, la a doua diviziune de clivaj, unul dintre blastomeri se împarte meridional și celălalt ecuatorial! Acest tip de clivaj se numește CLEAVAJ HOLOBLASTIC ROTAȚIONAL.
În plus, planurile de clivaj sunt oarecum diferite de cele ale altor animale. Primul clivaj este meridional, la fel ca în cazul ariciului de mare și al broaștei. Cu toate acestea, la a doua diviziune de clivaj, unul dintre blastomeri se împarte meridional și celălalt ecuatorial! Acest tip de clivaj se numește CLEAVAJ HOLOBLASTIC ROTAȚIONAL.
O altă caracteristică unică a clivajului mamiferelor este că clivajele blastomerilor sunt asincrone. (în comparație cu sincronia ariciului de mare și a broaștei până la tranziția de la jumătatea blastulei). Clivajul embrionului de mamifere este reglat de nucleul ziotic încă de la început.
Prin cel de-al treilea clivaj, blastomerii formează un ghem de celule slab asociate, la fel ca la celelalte animale pe care le-am studiat. Înaintea celei de-a patra clivaje, celulele din blastulă își schimbă dramatic comportamentul unele față de altele. Acestea încearcă rapid să își maximizeze contactele cu ceilalți blastomeri și, prin aceasta, determină compactarea blastulei.

Această COMPACTIUNE rezultă în parte din producerea unei noi molecule de adeziune UVOMORULIN (E-Cadherin) și este stabilizată prin formarea de joncțiuni strânse între celulele exterioare care, ca la ariciul de mare, sigilează interiorul blastulei de exterior. De asemenea, celulele formează între ele joncțiuni lacunare care permit trecerea moleculelor mici, cum ar fi ionii și unele molecule de mesager secundar, cum ar fi Ca++ și C-AMP. Morula compactă cu 16 celule este formată dintr-o crustă exterioară de celule și câteva celule (1-2) complet interne. Cele mai multe dintre celulele externe dau naștere CELULELOR TROBLASTICE SAU TROFECTODERMICE. Aceste celule nu contribuie la embrionul propriu-zis, dar în schimb sunt necesare pentru implantarea embrionului în peretele uterin și formează țesuturile CHORIANULUI, o componentă esențială a placentei despre care vom vorbi mai târziu.
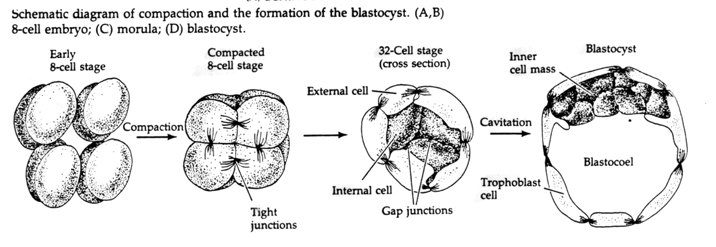
Celulele embrionului derivă din cele câteva celule interne ale blastulei în stadiu de 16 celule. Aceste celule generează masa celulară interioară de celule din care se dezvoltă întregul embrion. Până la cea de-a 6-a clivaj, stadiul de 64 de celule, masa celulară internă și stratul trofoblastic sunt complet separate. Trofoblastele secretă lichid în interior pentru a crea blastocoelul. Embrionul se numește acum blastocist.
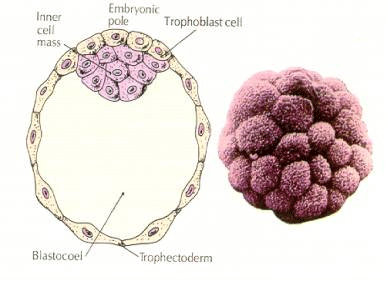 FORMAȚIA MASA CELULARĂ INTERIOARĂ
FORMAȚIA MASA CELULARĂ INTERIOARĂ
Cum sunt create aceste celule din masa celulară interioară? Există anumiți blastomeri predestinați prin factori intrinseci să devină progenitori ai masei celulare interne? Răspunsul pare să fie negativ. Toți blastomerii timpurii par a fi totipotenți, iar determinarea celulelor care vor contribui la stratul trofoblastic și care la masa celulară internă este pur și simplu o chestiune de poziție întâmplătoare. Celulele de la un embrion în stadiu de 4 celule, care în mod normal vor da naștere atât la masa celulară internă, cât și la celule de trofectoderm, transplantate în exteriorul unui embrion în stadiu de 32 de celule, dau naștere doar la trofectoderm. Acestea nu contribuie la formarea embrionului propriu-zis. Amintiți-vă din prelegerea anterioară despre clonare că fuziunea a doi embrioni de șoarece în stadiul de 8 celule are ca rezultat un embrion normal, ceea ce sugerează că toate celulele din acel stadiu sunt totipotente.
CLIVAJUL MEROBLASTIC
În ouăle telolecitale și centrolecitale, gălbenușul mare și dens împiedică clivajul. Ouăle telolecitale sunt caracteristice păsărilor, peștilor și reptilelor, în timp ce ouăle centrolecitale sunt caracteristice insectelor. Ouăle telolecitale au ca rezultat un clivaj discoidal meroblastic. Clivajul se limitează la blastodiscul de la polul animal al oului. La clivajele timpurii, deoarece clivajul nu poate trece prin gălbenuș, blastomerii sunt continui la marginile lor vegetale.
Acest film despre dezvoltarea peștelui zebră realizat deRolf Karlstrom este excelent. (Film de Paul Myers)
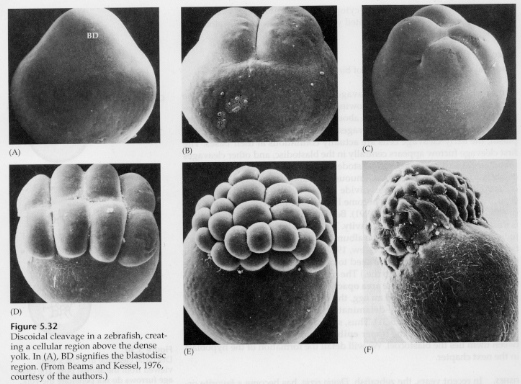 Nu înainte de clivajele ecuatoriale celulele blastodermului se separă de gălbenuș. Alte clivaje ecuatoriale creează un blastoderm multistratificat cu o grosime de trei sau patru celule.
Nu înainte de clivajele ecuatoriale celulele blastodermului se separă de gălbenuș. Alte clivaje ecuatoriale creează un blastoderm multistratificat cu o grosime de trei sau patru celule.
La păsări se formează un spațiu între blastoderm și gălbenuș numit cavitatea SUBGERMINALĂ. Până la diviziunea 16 (60.000 de celule) celulele din blastoderm migrează în cavitatea subgerminală pentru a forma un al doilea strat. Cele două straturi se numesc EPIBLAST extern și HIPOBLAST intern, cu blastocelul între ele. Vom studia acest lucru mai în detaliu mai târziu, când vom discuta despre gastrularea păsărilor și a mamiferelor
Oualele de artropode suferă un CLEAVAJ SUPERFICIAL. Masa mare centrală mare de gălbenuș limitează clivajul la marginea citoplasmatică a oului.
O variație interesantă și informativă este observată la insecte. Nucleii zigotici se divid fără clivaj. Adică nucleele suferă cariokineza—-diviziunea mitotică a nucleului— fără citokineză—diviziunea celulei. Aceste nuclee goale se numesc ENERGII. Nucleii se divid cu o viteză uimitoare – la fiecare 8 minute (toată embriogeneza durează doar 22 de ore).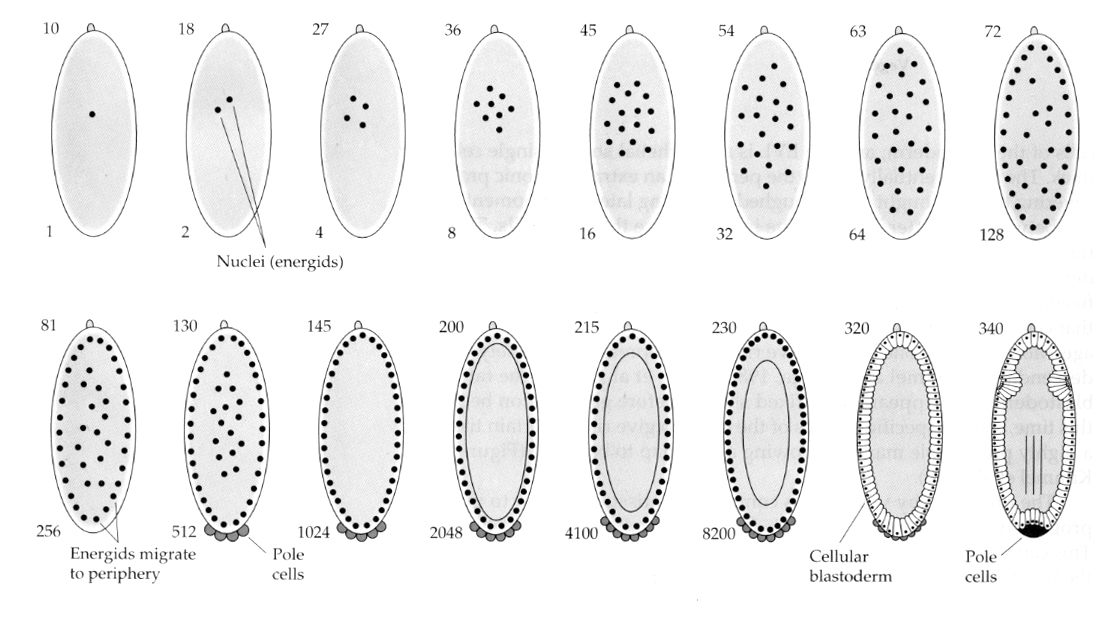
 După mai multe runde de cariocineză, nucleii goi migrează la periferia oului. În acest stadiu se numește BLASTODERM SINCITAL pentru că toți nucleii împart aceeași citoplasmă. Celularizarea are loc în jurul celei de-a 14-a diviziuni nucleare pentru a crea BLASTODERMUL CELULAR. După acest moment, celulele se divid în mod asincron. Acest lucru corespunde tranziției midblastula a broaștelor și aricii de mare. (trecerea de la expresia genei materne la expresia genetică preponderent zigottică) Amintiți-vă că s-a considerat că tranziția midblastula a fost declanșată de raportul dintre cromatină și citoplasmă. Dovezile pentru acest mecanism la muște sunt observate prin examinarea embrionilor haploizi mutanți. Acești embrioni suferă tranziția midblastula și celulizarea o diviziune mai târziu 15. În plus, puteți accelera celulizarea prin legarea oului și reducerea volumului de citoplasmă. Deși stadiul de blastoderm sincițial sugerează că toți nucleii sunt echipotenți, în sensul că nu par să existe bariere difuzionale pentru determinanții citoplasmatici, de fapt citoplasma este foarte regionalizată, iar nucleii au în jurul lor domenii citoplasmatice foarte bine organizate.
După mai multe runde de cariocineză, nucleii goi migrează la periferia oului. În acest stadiu se numește BLASTODERM SINCITAL pentru că toți nucleii împart aceeași citoplasmă. Celularizarea are loc în jurul celei de-a 14-a diviziuni nucleare pentru a crea BLASTODERMUL CELULAR. După acest moment, celulele se divid în mod asincron. Acest lucru corespunde tranziției midblastula a broaștelor și aricii de mare. (trecerea de la expresia genei materne la expresia genetică preponderent zigottică) Amintiți-vă că s-a considerat că tranziția midblastula a fost declanșată de raportul dintre cromatină și citoplasmă. Dovezile pentru acest mecanism la muște sunt observate prin examinarea embrionilor haploizi mutanți. Acești embrioni suferă tranziția midblastula și celulizarea o diviziune mai târziu 15. În plus, puteți accelera celulizarea prin legarea oului și reducerea volumului de citoplasmă. Deși stadiul de blastoderm sincițial sugerează că toți nucleii sunt echipotenți, în sensul că nu par să existe bariere difuzionale pentru determinanții citoplasmatici, de fapt citoplasma este foarte regionalizată, iar nucleii au în jurul lor domenii citoplasmatice foarte bine organizate.
MECANISMELE CLEAVAJULUI
Ciclul celular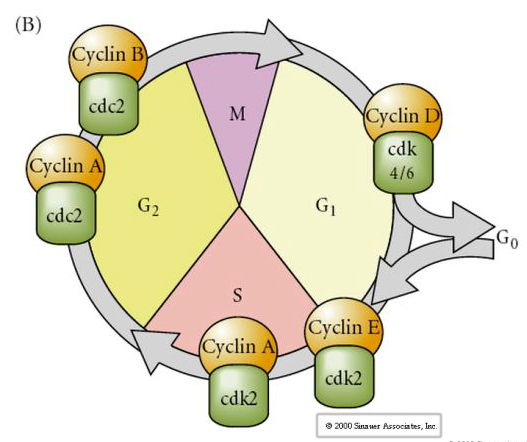
M-mitoză
G1- decalaj de pre-replicare
S- sinteză de ADN
G2-decalaj premitotic
În embrionii în stadiul de clivaj, cum ar fi broaștele și muștele, blastomerii trec direct de la M la S, fără a interveni în stadiile G1 sau G2. După tranziția midblastula celulele din ambele animale au un G1 și G2. Experimentele de transplant elegant au demonstrat că citoplasma este cea care reglează atât cariochineza, cât și citochineza. Dacă nuclee din celule în diviziune sunt transplantate în ovocite, acestea se opresc imediat din diviziune.
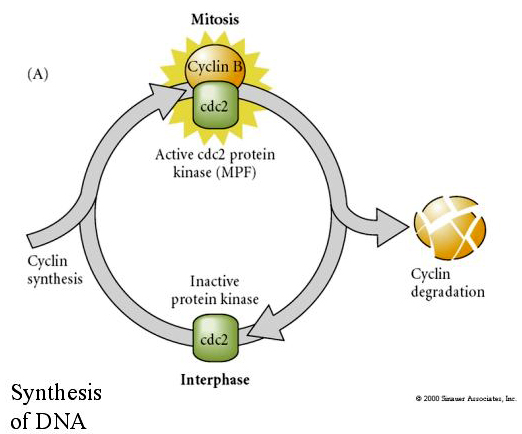 În schimb, dacă nuclee din celule care nu se divid sunt puse în ovule enucleate fertilizate, acestea încep să se dividă. Ovulele enucleate activate artificial fără centrioli vor suferi contracții corticale care amintesc de clivaj. Au fost identificați unii dintre factorii citoplasmatici care reglează diviziunea celulară în embrionul timpuriu.
În schimb, dacă nuclee din celule care nu se divid sunt puse în ovule enucleate fertilizate, acestea încep să se dividă. Ovulele enucleate activate artificial fără centrioli vor suferi contracții corticale care amintesc de clivaj. Au fost identificați unii dintre factorii citoplasmatici care reglează diviziunea celulară în embrionul timpuriu.
FACTORUL CITOSTATIC (CSF) este ridicat după prima diviziune meiotică și oprește ovocitele în a doua metafază meiotică. La fecundare, Ca inactivează CSF, meioza este finalizată și pronucleii fuzionează.
FACTORUL DE PROMOVARE A MITOSISULUI (MPF) determină celulele să intre în faza M. Activarea MPF determină: 1. condensarea cromozomilor prin fosforilarea histonei H1, 2. descompunerea învelișului nuclear prin hiperfosforilarea celor 3 lamine nucleare, 3. inhibarea ARN polimerazei pentru a opri transcripția, 4. fosforilarea subunității de reglare a miozinei pentru a inhiba citochineza.
Model sugerat pentru reglarea ciclică a ciclului celular în timpul etapelor de clivaj ale embrogenezei. MPF induce celula să treacă de la S la M. CSF se leagă de MPF și împiedică inactivarea sa. Celula rămâne în M. Ca crește și determină inactivarea CSF care, la rândul său, duce la inactivarea MPF, iar celula trece din M în S și ciclul se repetă. MPF este alcătuit din două subunități, ciclina B și cdc2. Ciclina B este cea care suferă o sinteză și o degradare specifice ciclului celular, reglementate de nucleul celulelor pentru a controla ciclul celular în celulele somatice normale. Cu toate acestea, în timpul oogenezei, ovulul este încărcat cu „regulatori” ai ciclinei B și ARNm al ciclinei B, astfel încât sinteza acesteia este reglată de factori materni independenți de nucleul zigotului. Astfel, abia după ce componentele materne „se epuizează”, nucleul zigottic preia controlul și se revine la un ciclu celular normal (M, G1, S, G2). 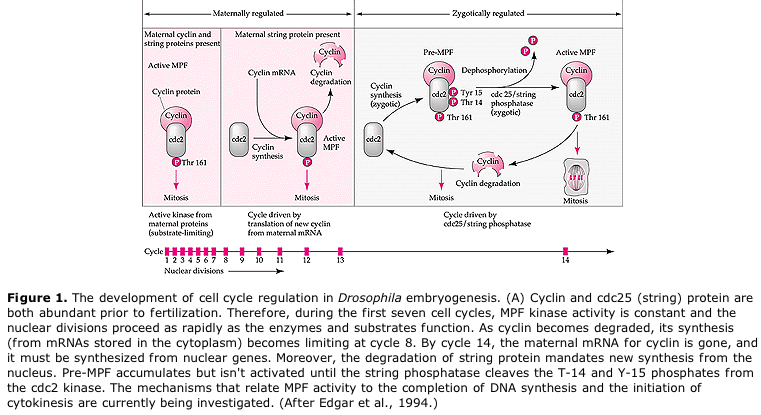
DETERMINAREA FAȚEI CELULELOR
Localizarea citoplasmatică a DETERMINANȚILOR ca mecanism general și de bază pentru structurarea timpurie (exemple Tunicate și arici de mare). O întrebare majoră a biologiei dezvoltării este când și cum se determină destinele celulare în timpul dezvoltării. Aceasta este strâns legată de întrebarea privind modul în care are loc formarea modelelor în timpul dezvoltării. Embrionul nu numai că trebuie să genereze numărul și tipul corect de celule diferențiate, dar acestea trebuie să fie organizate în mod corect în raport cu toate celelalte celule din embrion pentru a forma un animal funcțional. Vom examina două posibilități de determinare a destinului celular și de formare a tiparelor: 1. 1. Soarta celulară ar putea fi determinată de factori intrinseci introduși în ovul în timpul oogenezei și apoi repartizați în blastomeri specifici în timpul clivajului; 2. Semnalele extrinseci furnizate de mediul embrionar ar putea furniza informațiile de modelare pentru a regla soarta celulară. După cum vom vedea, majoritatea organismelor complexe utilizează o combinație de semnale intrinseci și extrinseci pentru a regla destinul celular și formarea modelului embrionar.
Specificarea autonomă a destinului celular prin determinanți citoplasmatici sugerează că destinul unei celule depinde în întregime de linia sa genetică, în timp ce dezvoltarea „regulativă” sugerează că destinul unei celule este determinat de semnale externe de la alte celule. Aceste două mecanisme de specificare celulară pot fi distinse experimental prin experimente de izolare, ablație și transplant. Dacă un blastomer izolat dintr-un embrion se diferențiază în mod normal (ca și cum ar fi încă în poziția sa normală în embrion), putem spune că acesta trebuie să aibă determinanți intrinseci care îi specifică soarta. Cu toate acestea, dacă se diferențiază în mod anormal, putem spune că soarta sa celulară depinde de semnale externe. Dacă abatem un blastomer dintr-un embrion, iar embrionul se dezvoltă în mod anormal, lipsind toate destinele celulelor care apar în mod normal din blastomerul ablatat, vom spune că dezvoltarea este autonomă din punct de vedere celular și specificată intrinsec. Cu toate acestea, dacă embrionul se dezvoltă normal, spunem că blastomerii rămași își pot regla destinul celular pentru a compensa celulele lipsă. Dacă o celulă transplantată își menține soarta celulară pe baza poziției sale inițiale, atunci spunem că soarta sa a fost determinată, dacă ia o nouă soartă pe baza noii sale poziții transplantate, spunem că soarta sa este reglată de semnale externe de la celulele din apropiere.
Localizarea și reglarea citoplasmatică în ovulul tunicat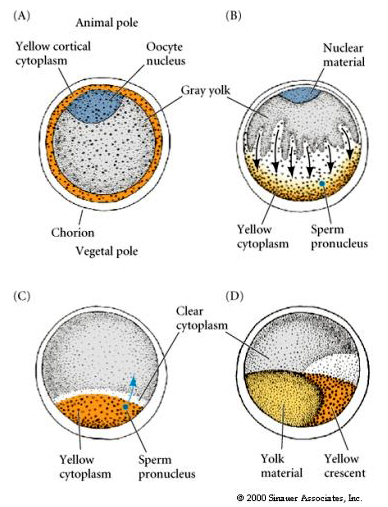
La sfârșitul oogenezei, ovulul tunicat are un pol animal și un pol vegetal clar distinse. Există o citoplasmă corticală galbenă care înconjoară o citoplasmă interioară gri gălbuie. Nucleul ovocitelor este deplasat spre polul animal. Intrarea spermatozoizilor în emisfera vegetală fertilizează ovulul și inițiază dezvoltarea. O rearanjare dramatică a citoplasmei ovulului are loc după fertilizare, dând naștere la citoplasme colorate regional, care par să se coreleze cu sorții blastomerilor subsecvenți.
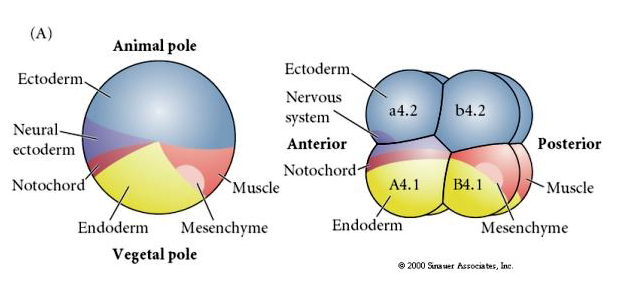 Rețineți că harta sorții se corelează cu citoplasmele de diferite culori ale embrionului tunicat. Nu vă lăsați confundați de culorile diferite din două figuri. Citoplasma galbenă „portocalie” în formă de semilună este corelată cu destinele musculare, iar citoplasma Yolky (galbenă) este corelată cu destinele endodermice. Citoplasma gri (albă sau purpurie albăstruie) de deasupra semilunii galbene este corelată cu ectodermul neural.
Rețineți că harta sorții se corelează cu citoplasmele de diferite culori ale embrionului tunicat. Nu vă lăsați confundați de culorile diferite din două figuri. Citoplasma galbenă „portocalie” în formă de semilună este corelată cu destinele musculare, iar citoplasma Yolky (galbenă) este corelată cu destinele endodermice. Citoplasma gri (albă sau purpurie albăstruie) de deasupra semilunii galbene este corelată cu ectodermul neural.
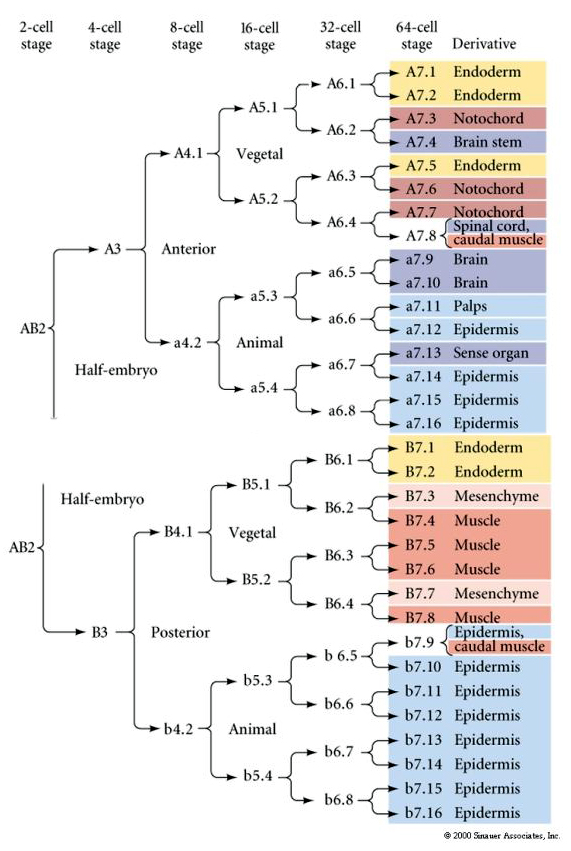 Această hartă a liniei de descendență arată corelația liniei de descendență invariante cu blastomere parcelate citoplasme particulare colorate de către clivajele celulelor invariante. Cu toate acestea, clivajele și liniile de linie invariante nu dovedesc neapărat o specificare autonomă a celulelor prin determinanți citoplasmatici.
Această hartă a liniei de descendență arată corelația liniei de descendență invariante cu blastomere parcelate citoplasme particulare colorate de către clivajele celulelor invariante. Cu toate acestea, clivajele și liniile de linie invariante nu dovedesc neapărat o specificare autonomă a celulelor prin determinanți citoplasmatici.
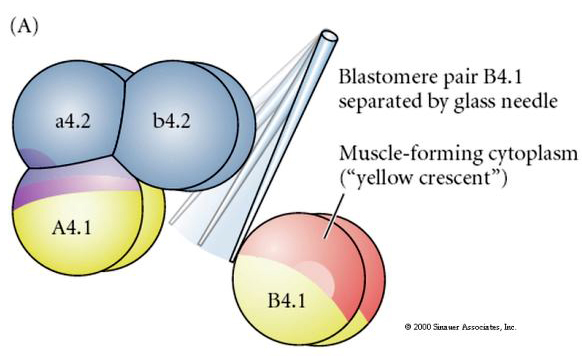 Sunt necesare manipulări experimentale pentru a testa determinarea regulativă versus cea autonomă a destinului celular. Experimentele clasice de izolare prezentate în următoarele trei figuri încearcă să demonstreze că destinul celular este determinat de determinanți citoplasmatici pe care aceștia îi dobândesc prin clivaje stereotipice. Se utilizează un ac de sticlă pentru a separa perechea de blastomeri B4.1 de restul embrionului. Blastomerii B4.1 dobândesc în mod normal citoplasma galbenă crecentă corelată cu destinul celulelor musculare.
Sunt necesare manipulări experimentale pentru a testa determinarea regulativă versus cea autonomă a destinului celular. Experimentele clasice de izolare prezentate în următoarele trei figuri încearcă să demonstreze că destinul celular este determinat de determinanți citoplasmatici pe care aceștia îi dobândesc prin clivaje stereotipice. Se utilizează un ac de sticlă pentru a separa perechea de blastomeri B4.1 de restul embrionului. Blastomerii B4.1 dobândesc în mod normal citoplasma galbenă crecentă corelată cu destinul celulelor musculare.
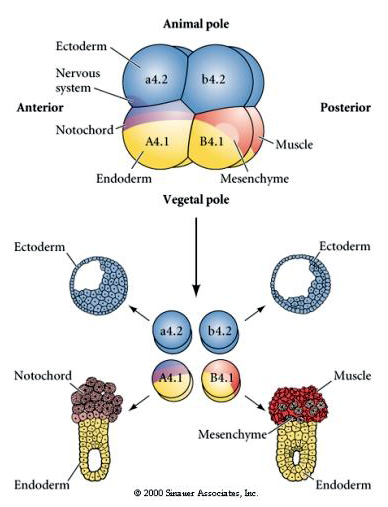
Aici se pot vedea rezultatele experimentelor de izolare. În fiecare caz, blastomerii izolați dau naștere doar acelui subset de destine celulare pe care le-ar produce în mod normal în embrionul intact. Blastomerii izolați nu își reglează destinul pentru a compensa pentru vecinii lor lipsă. Blastomerii de la polul animal, a4.2 și b4.2, dau naștere numai la celule ectodermice. A4.1 dă naștere la celule notocordale și endodermale, în timp ce B4.1 dă naștere la celule musculare și endodermale. Nici unul dintre blastomerii izolați nu poate da naștere la toate componentele celulare ale unui embrion normal.
În următorul experiment de mai jos se folosește un ac pentru a manipula planul de clivaj ecuatorial astfel încât acesta să fie mai vegetal decât în mod normal și acum blastomerii polului animal, b4.2, dobândesc o parte din citoplasma „semilună galbenă”. Atunci când acești blastomeri sunt izolați, ei dau acum naștere unor celule musculare. Acest lucru demonstrează foarte bine că citoplasma „semilună galbenă” poate determina destinul celulelor musculare și poate face acest lucru într-o manieră autonomă din punct de vedere celular.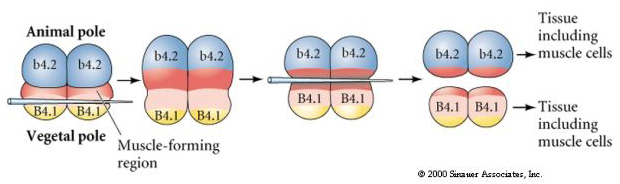
LOCALIZARE ȘI REGLEMENTARE ÎN OUL DE URSINĂ MARE
Un canal gelatinos definește localizarea polului animal și reflectă polaritatea timpurie a oului. Modelul timpuriu al clivajelor nu depinde de locul de intrare a spermatozoizilor, ci sunt determinate de polaritatea intrinsecă/asimetria oului. Boveri (1901) a descris o bandă subecuatorială de pigment dispusă ortongonal față de axa animal-vegetal. Acești granule indicau, de asemenea, localizarea citoplasmei care este inclusă mai târziu în celulele arheenteronului. Horstadius (1928) a separat blastomerele animale și vegetale și a arătat că numai blastomerele vegetale vor da naștere la micromere, vor fi gastrulate și vor forma scheletul. Concluzia sa a fost că factorii citoplasmatici localizați în jumătatea vegetală sunt necesari pentru micromere, gastrulare și formarea arcteronului, precum și pentru formarea scheletului.Rețineți modelul clivajelor timpurii. Micromerele apar în timpul celei de-a patra clivaje (stadiul de 16 celule) dintr-o diviziune ecuatorială inegală a blastomerilor de la polul vegetal.
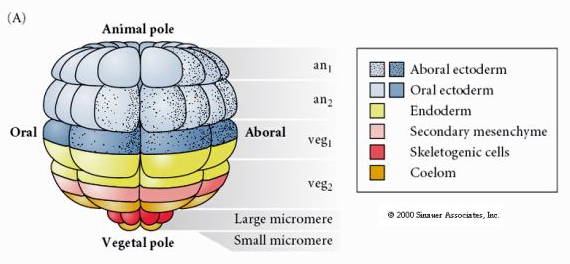 Aceasta arată harta destinului blastulei de arici de mare în stadiul de 64 de celule. Observați că micromerele sunt celulele primare ale mezenchimului și dau naștere scheletului larvar (spiculele din stadiul de pluteu).
Aceasta arată harta destinului blastulei de arici de mare în stadiul de 64 de celule. Observați că micromerele sunt celulele primare ale mezenchimului și dau naștere scheletului larvar (spiculele din stadiul de pluteu).
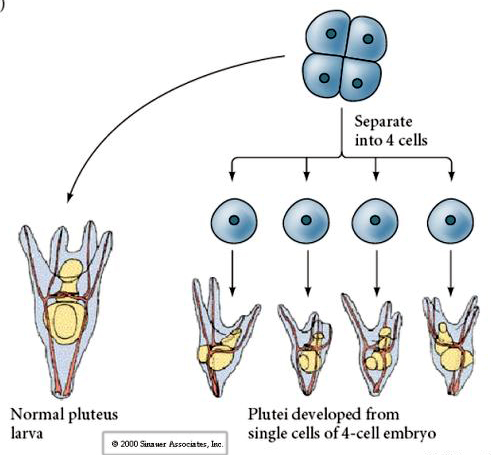 În stadiul de patru celule, dacă blastomerele sunt izolate unele de altele, ele sunt capabile să își „regleze” soarta și să dea naștere la 4 larve mici din stadiul de pluteu.
În stadiul de patru celule, dacă blastomerele sunt izolate unele de altele, ele sunt capabile să își „regleze” soarta și să dea naștere la 4 larve mici din stadiul de pluteu.
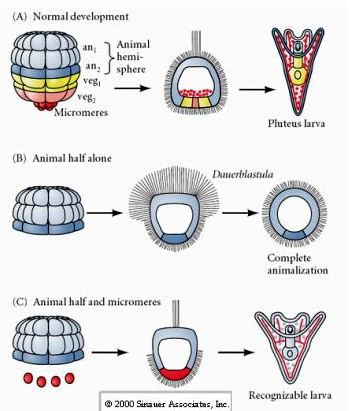 În schimb, în stadiile ulterioare, dacă se izolează jumătăți de blastomere animale, se constată că acestea produc doar o dauerblastulă „animalizată” care nu exprimă nici un destin celular mezodermal sau endodermal. Jumătate de blastomeri vegetali izolați dau naștere la larve care exprimă sorți celulari ectodermici, mezodermici și endodermici, ceea ce arată că soarta acestor celule poate fi reglată. Micromerele izolate (mezenchimul primar) sunt supuse unui număr corect de diviziuni celulare și dau naștere ÎNTOTDEAUNA la spiculețe conform programului. Astfel, micromerele sunt specificate definitiv ca fiind precursoare ale celulelor mezenchimului scheletic atunci când apar pentru prima dată în stadiul de 16 celule. Experimentele cheie au fost punerea în comun a micromerelor cu blastomerele de la polul animal și demonstrarea faptului că, deși soarta micromerelor a fost „fixată sau determinată” în momentul nașterii lor, micromerele au fost capabile să „inducă” noi sorți celulare în blastomerele de la polul animal. Micromerele au fost capabile de a induce sorții endodermice și mezodermice în blastomerii de la polul animal! Astfel, ultimul experiment din „C” arată că, atunci când se adaugă micromeri la o jumătate de blastulă a polului animal, acum se poate induce formarea unei larve recognoscibile, care să exprime sorți endodermici, mezodermici și ectodermici.
În schimb, în stadiile ulterioare, dacă se izolează jumătăți de blastomere animale, se constată că acestea produc doar o dauerblastulă „animalizată” care nu exprimă nici un destin celular mezodermal sau endodermal. Jumătate de blastomeri vegetali izolați dau naștere la larve care exprimă sorți celulari ectodermici, mezodermici și endodermici, ceea ce arată că soarta acestor celule poate fi reglată. Micromerele izolate (mezenchimul primar) sunt supuse unui număr corect de diviziuni celulare și dau naștere ÎNTOTDEAUNA la spiculețe conform programului. Astfel, micromerele sunt specificate definitiv ca fiind precursoare ale celulelor mezenchimului scheletic atunci când apar pentru prima dată în stadiul de 16 celule. Experimentele cheie au fost punerea în comun a micromerelor cu blastomerele de la polul animal și demonstrarea faptului că, deși soarta micromerelor a fost „fixată sau determinată” în momentul nașterii lor, micromerele au fost capabile să „inducă” noi sorți celulare în blastomerele de la polul animal. Micromerele au fost capabile de a induce sorții endodermice și mezodermice în blastomerii de la polul animal! Astfel, ultimul experiment din „C” arată că, atunci când se adaugă micromeri la o jumătate de blastulă a polului animal, acum se poate induce formarea unei larve recognoscibile, care să exprime sorți endodermici, mezodermici și ectodermici.
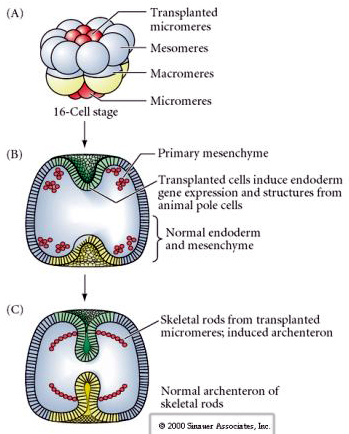 Ultimul set de experimente demonstrează că, chiar și la un embrion normal, dacă se transplantează micromeri la calota polului animal, se poate induce un arpenteron secundar și se poate modifica structura axială normală. Acest lucru argumentează din nou că micromerele dobândesc un derminant citoplasmatic care le specifică destinul celular și că ele furnizează semnalul inductiv care modelează structurile axiale ale embrionului de uchin de mare. Soarta micromerelor nu poate fi modificată, dar semnalele de la micromere pot modifica soarta tuturor celorlalți blastomeri.
Ultimul set de experimente demonstrează că, chiar și la un embrion normal, dacă se transplantează micromeri la calota polului animal, se poate induce un arpenteron secundar și se poate modifica structura axială normală. Acest lucru argumentează din nou că micromerele dobândesc un derminant citoplasmatic care le specifică destinul celular și că ele furnizează semnalul inductiv care modelează structurile axiale ale embrionului de uchin de mare. Soarta micromerelor nu poate fi modificată, dar semnalele de la micromere pot modifica soarta tuturor celorlalți blastomeri.
Horstadius: (1928, 1935) a arătat experimental că, într-un embrion în stadiu de 16 celule, toate etajele de blastomeri, cu excepția micromerelor, vor lua destine diferite atunci când sunt transplantate în poziții diferite în embrioni chimeri. Arheenteronul se va dezvolta din blastomeri veg 1 dacă celulele veg 2 sunt îndepărtate, iar micromerii sunt plasați în contact cu stratul veg 1. În absența micromerelor, blastomerii veg 2 dau naștere la archenteron și la structurile scheletice. În mod clasic, pentru a explica aceste rezultate, a fost invocat un gradient dual animal-vegetal. Cu toate acestea, aceste rezultate indică doar faptul că interacțiunile inductive decisive au loc între nivelurile de blastomeri adiacenți.
Micromerele individuale implantate în apropierea polului animal inhibă formarea de tufișuri apicale și, în unele cazuri, induc o nouă axă embrionară. Blastomerele Veg 2 vor induce, de asemenea, modificări similare cu cele ale micromerelor atunci când sunt transplantate lângă blastomerele de la polul animal.
REZULTATUL GENERAL AL TRANSPLANTAȚIILOR: se constată că soarta unor blastomere date este întotdeauna afectată de apoziția diferitelor celule vecine care le sunt adiacente în embrionii normali.
HIPOTEZĂ: Determinanții citoplasmatici materni localizați specifică anumite celule din embrionul normal, în special micromerele și precursorii arheteronului din apropierea polului vegetal. Aceste celule determină apoi în mod inductiv destinele blastomerilor vecini, care interacționează la rândul lor cu vecinii lor. Mulți dintre blastomeri păstrează alte potențialități decât cele pe care le exprimă în mod normal, iar pentru o anumită perioadă de timp acești blastomeri sunt specificați doar în mod reversibil, așa cum este necesar pentru un sistem de dezvoltare care depinde în mare măsură de inducție.