Cleavage viittaa stereotyyppiseen varhaisen mitoottisen jakautumisen malliin, joka jakaa suuren määrän munasoluplasmaa. Varhainen zygootti on ainutlaatuinen, koska se on niin suuri. Useimmat solut kasvavat mitoosijaksojen välillä, mutta tämä ei päde varhaisen pilkkoutumisvaiheen blastomeereihin. Jokaisen jakautumisen myötä solut pienenevät. Tämä solujen nopea jakautuminen ilman samanaikaista kasvua pysähtyy äkillisesti vaiheessa, jota kutsutaan keskiblastomeren siirtymävaiheeksi ja jossa zygootin ydin ottaa solusyklin hallintaansa.
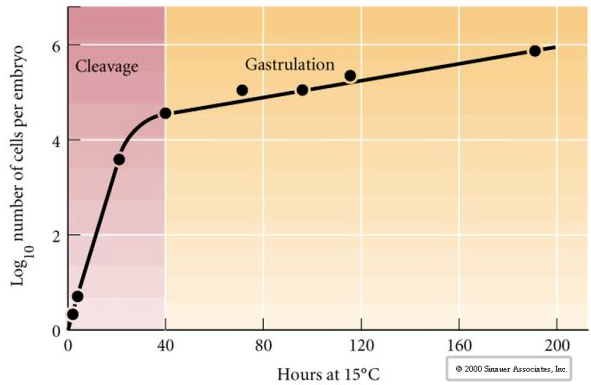 On olemassa viitteitä siitä, että jokin äidin tekijä, kenties transkriptionaalinen säätelijä, on vastuussa tästä varhaisesta nopeasta solunjakautumismallista. Muuttamalla keinotekoisesti sytoplasman ja ydin-DNA:n suhdetta voidaan muuttaa keskirakkulan siirtymän ajankohtaa. Midblastula-siirtymällä tarkoitetaan ajankohtaa, jolloin tapahtuu merkittävä siirtymä äidin geenien ilmentymisestä zygoottisten geenien ilmentymiseen.
On olemassa viitteitä siitä, että jokin äidin tekijä, kenties transkriptionaalinen säätelijä, on vastuussa tästä varhaisesta nopeasta solunjakautumismallista. Muuttamalla keinotekoisesti sytoplasman ja ydin-DNA:n suhdetta voidaan muuttaa keskirakkulan siirtymän ajankohtaa. Midblastula-siirtymällä tarkoitetaan ajankohtaa, jolloin tapahtuu merkittävä siirtymä äidin geenien ilmentymisestä zygoottisten geenien ilmentymiseen.
Joidenkin lajien hedelmöittyminen johtaa radikaaleihin sytoplasman liikkeisiin, jotka ovat välttämättömiä sen varmistamiseksi, että sytoplasman determinantit sijaitsevat oikeissa paikoissa suhteessa myöhempiin pilkkoutumistapahtumiin.
EMBRYONISEN HAJAUTUMISEN PATTERIT
Embryonisen hajautumisen malli määräytyy sekä mitoottisten spindeleiden sijainnin että keltarauhasen määrän ja jakautumisen mukaan. Keltuainen pyrkii estämään pilkkoutumista. Se hidastaa sitä tai jopa estää täydellisen pilkkoutumisen. Keltuainen on sopeutumista eläimille, jotka käyvät läpi enemmän tai vähemmän alkionmuodostuksen eristyksissä kaikesta ravinnosta. Joillakin eläimillä, kuten merisiilillä, on suhteellisen vähän keltuaista, koska ne kehittyvät nopeasti vapaasti uivaksi toukkamuodoksi, joka saa ravintoa ympäristöstään. Toiset eläimet, kuten pussieläimet, syntyvät ennenaikaisesti, mutta saavat ravintoa vanhempien pussissa. Istukkanisäkkäät kehittävät erikoistuneen elimen, jonka kautta alkio saa ravintoa koko kehityksensä ajan, joten niillä on myös vähän keltuaista.
Munien tyypit keltuaisten ominaisuuksien perusteella kuvataan seuraavasti:
Isolecithal: niukasti tasaisesti jakautunutta keltuaista, esim. merisiili, hiiri
Mesolecthal: kohtuullisen paljon keltuaista, usein epätasaisesti jakautunutta, esim, sammakko
Telolecithal: tiheä keltuainen keskittynyt toiseen päähän, esim. lintu, matelija
Centrolecithal: keltuainen keskittynyt munan keskelle, esim. kärpänen
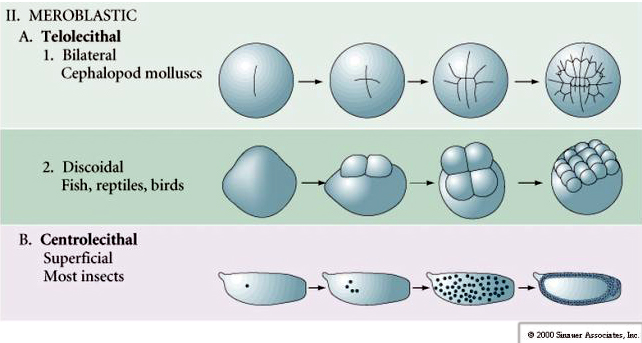
Monet munat ovat polarisoituneita siten, että niissä on keltuaista runsaasti sisältävä napa, jota kutsutaan kasvinapaksi, ja keltuaista köyhä napa, jota kutsutaan eläinnapaksi, esim. sammakko. Zygoottiytimen ydin on yleensä siirtynyt kohti eläimellistä napaa. Zygootit, joissa on suhteellisen vähän keltuaista (isolecithal ja mesolecithal), pilkkoutuvat HOLOBLASTISESTI. Halkaisukouru ulottuu koko munan läpi. Telolecithaaliset ja centrolecithaaliset zygootit puolestaan halkeavat MEROBLASTISESTI, jolloin halkaisutaso ulottuu vain keltuaiseen asti. Sentrolecithal-munissa (monet hyönteisten munat) halkio on meroblastinen ja pinnallinen, kun taas telolecithal-munissa (linnut ja kalat) halkio on diskoidinen
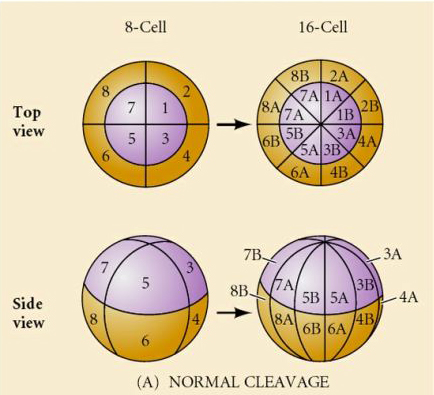
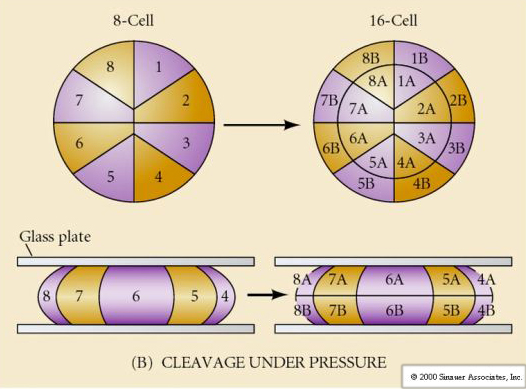
Luonnossa nähdään useita erilaisia halkion symmetriatyyppejä: radiaalinen (piikkinahkaiset, sammakkoeläimet), spiraalinen (nilviäiset, rengaskuoriaiset), bilateraalinen (ascidiat, sädekehäeläimet, tunikoidit), rotaatiomallia oleva symmetriaa noudattava symmetriaa noudattava symmetriaa noudattava symmetriaa noudattava symmetriaa noudattava symmetriaa noudattava symmetria. Alla olevissa kahdessa kuvassa on esimerkkejä holoblastisesta ja meroblastisesta pilkkoutumissymmetriasta.
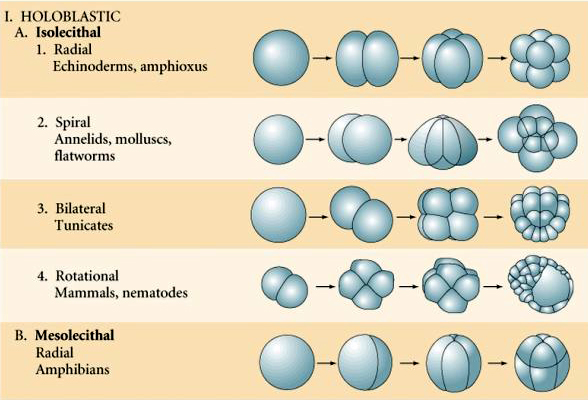
RADIAALINEN HOLOBLASTINEN PURISTUMINEN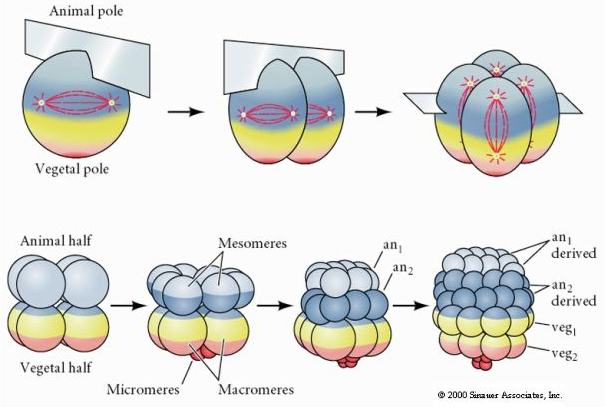
Erinomainen filmi merisiilin pilkkoutumisesta Rachel Finkin teoksesta ”A Dozen Eggs”.
Merisiileillä on myös radiaalinen holoblastinen pilkkoutuminen, mutta niillä on joitakin mielenkiintoisia eroja. Ensimmäinen halkio on meridionaalinen.Toinen halkio on meridionaalinen. Kolmas pilkkoutuminen on ekvatoriaalista Neljäs pilkkoutuminen on meridiaalista, mutta siinä missä neljä eläimen napaan kuuluvaa solua jakautuu tasaisesti synnyttäen kahdeksan samankokoista eläimen blastomeeria, joita kutsutaan MESOMEREIKSI, kasvisolujen solut jakautuvat epäsymmetrisesti ekvatoriaalista tasoa pitkin synnyttäen neljä isoa MACROMERIA ja neljä paljon pienempää MICROMERIA kasvin napaan. Viidennessä jakautumisessa MESOMERES jakautuu ekvatoriaalisesti muodostaen kaksi kahdeksan MESOMERESin kerrosta an1 ja an2 , MACROMERES jakautuu meridiaalisesti muodostaen kahdeksan solun kerroksen an2:n alapuolella, MICROMERES jakautuu muodostaen veg1-kerroksen alapuolella olevan solurykelmän. Kuudennet jakautumat ovat kaikki ekvatoriaalisia, jolloin syntyy veg2-kerros. Seitsemäs jakautuminen on meridonaalista, jolloin syntyy 128-soluinen blastula.
Miten nämä jakautumismallit määräytyvät? Ovatko ne riippuvaisia edellisestä jakautumisesta ja toistetaanko ne kuin nauhalta vai määrääkö ne jokin sisäinen kello? Vuonna 1939 Horstadius esti yhden tai kaksi kolmesta ensimmäisestä pilkkoutumisesta ja havaitsi, että mikromereenien ilmaantuminen tapahtui oikeaan aikaan riippumatta pilkkoutumishistoriasta
Johtopäätös näistä kokeista on, että munasolun kasvullisessa napassa on jokin tekijä, joka määrää mikromereenien muodostumisen, ja edelleen, että on oltava olemassa molekulaarinen kello, joka käynnistyy munasolun aktivoinnista. Kello on riippumaton varsinaisesta pilkkoutumistapahtumasta.
128-soluinen blastula on melko löysä solupallo, joka ympäröi onttoa blastokuorta. Pallo on yhden solukerroksen paksuinen, ja kaikki solut ovat kosketuksissa ulkoisen hyaliinikerroksen ja blastokotelon sisäisen nesteen kanssa. Tässä kehitysvaiheessa solut alkavat muodostaa epiteelille ominaisia tiiviitä liitoksia. Keskimmäinen blastokotelo on nyt eristetty ulkoisesta ympäristöstä. Blastomeerit jatkavat jakautumistaan akselinsa suuntaisesti hyaliinikerrokseen nähden, jolloin ne jäävät yhden solun paksuiseksi epiteeliksi. Blastokysta jatkaa laajentumistaan.
Kahdella teorialla yritetään selittää blastokystan laajentumismallia
1. Osmoottisen teorian mukaan blastomeereistä erittyy ioneja ja proteiineja blastokoteloon, mikä johtaa veden osmoottisen virtauksen aiheuttamaan paineen nousuun. Tämä paine olisi sitten vastuussa blastomeerien mitoosiakselin suuntaamisesta ja blastokotelon laajentumisesta.
2. Wolpertin ja hänen kollegoidensa esittämässä vaihtoehtoisessa teoriassa ehdotetaan, että blastomeerien väliset sekä blastomeerien ja hyaliinikerroksen väliset adhesiiviset vuorovaikutussuhteet todella suuntaavat mitoosiakselit. Toisin sanoen adheesio hyaliinikerrokseen on suurin, adheesio muihin blastomeereihin on seuraava, ja lopuksi vuorovaikutus blastokotelon seinämän kanssa on vähäisin. Eläimen navan kohdalla ensimmäinen pilkkoutuminen etenee noin 1 mm/min, kun taas kasvinavan kohdalla se etenee 50-100 kertaa hitaammin (.02 mm/min). Kun ensimmäinen pilkkoutuminen on vielä epätäydellistä munan keltakasvualueella, toinen meridionaalinen pilkkoutuminen alkaa tapahtua.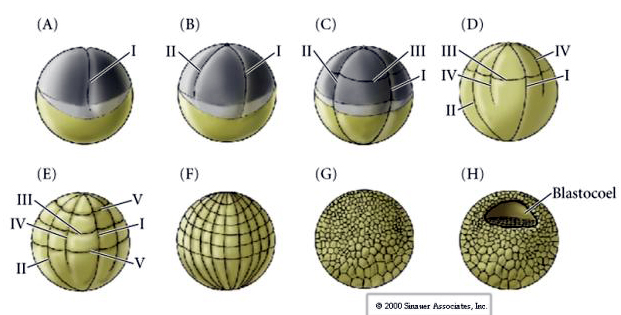
Kolmas pilkkoutuminen on ekvatoriaalista, mutta koska ytimet ja asterit ovat siirtyneet eläimen suuntaan, pilkkoutumistaso, vaikka se on kohtisuorassa eläinkasvualueen akseliin nähden, on myös siirtynyt eläinnapaan päin eikä jaa blastomeerejä tasaisesti. Tuloksena on neljä pienempää eläinblastomeeria (ns. MICROMERES) ja neljä suurta kasvin napaan liittyvää blastomeeria (ns. MACROMERES). Tämä epätasainen holoblastinen jakautuminen synnyttää nopeammin jakautuvan eläinnavan, joka koostuu pienemmistä mikromereistä, ja hitaammin jakautuvan kasvinnavan, joka koostuu makromereistä. Eläinnapa koostuu pian monista pienistä mikromereistä ja kasvinnapa muutamasta keltuaisen täyttämästä suuresta makromereistä. Vaikka blastokotelon muodostuminen alkaa ensimmäisestä jakautumisesta, se tulee ilmeiseksi vasta 128 solun vaiheessa.
MITÄ FUNKTIOITA BLASTOKOTELO PALVELEE?
Blastokotelo erottaa solut toisistaan avaruudellisesti niin, etteivät ne kosketa toisiaan. Blastokotelon katolla olevista soluista tulee tavallisesti ektodermiä. Jos blastokotelon katon soluja siirretään blastokotelon tyvessä olevien keltasolujen viereen, ne erilaistuvat mesodermiksi. Mesodermijohdokset syntyvät tavallisesti endodermien esiasteiden vieressä olevista soluista. Yksi mahdollisuus, jota aiomme tutkia perusteellisesti, on se, että kasvulliset solut saavat solusolujen vuorovaikutuksen kautta viereiset solut muuttumaan mesodermisiksi. Näin ollen blastokotelon muodostuminen voi olla välttämätöntä, jotta estetään epätarkoituksenmukaiset ”induktiiviset” vuorovaikutukset blastokystan varhaisten solujen välillä. Toinen ilmeinen blastokotelon tarve voi olla myöhemmässä kehitysvaiheessa, GASTRULAATIOSSA, jossa solut vaeltavat blastokotelon sisälle.
NAMMAISTEN MUNASOLU
Nisäkkäiden munasolu vapautuu munasarjasta munanjohtimeen, jossa se hedelmöittyy. Ensimmäinen pilkkoutuminen alkaa noin vuorokauden kuluttua hedelmöittymisestä munanjohtimessa. Jyrkässä ristiriidassa useimmista eläimistä nisäkkäiden hajaantuminen voi olla hyvin hidasta—1/vrk.
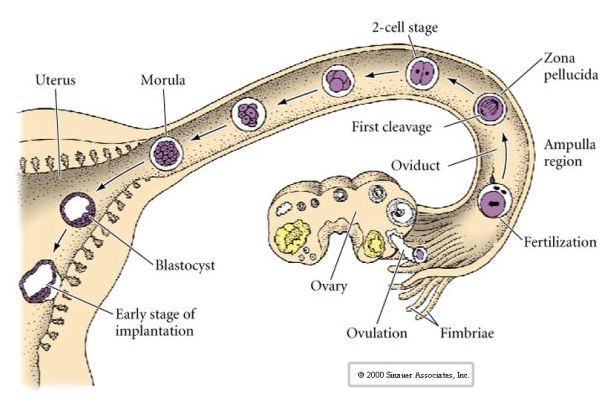
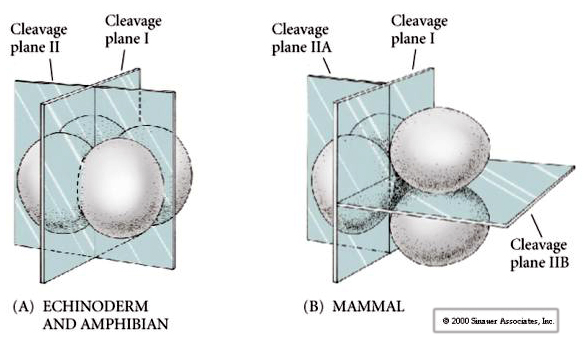 Lisäksi hajaantumistasot ovat hieman erilaiset kuin muilla eläimillä. Ensimmäinen halkio on meridiaalinen aivan kuten merisiilillä ja sammakolla. Toisessa pilkkoutumisessa toinen blastomeereistä jakautuu kuitenkin meridiaalisesti ja toinen ekvatoriaalisesti! Tällaista jakautumista kutsutaan ROTATIONAALISEKSI HOLOBLASTISEKSI HAJAUTUMISEKSI.
Lisäksi hajaantumistasot ovat hieman erilaiset kuin muilla eläimillä. Ensimmäinen halkio on meridiaalinen aivan kuten merisiilillä ja sammakolla. Toisessa pilkkoutumisessa toinen blastomeereistä jakautuu kuitenkin meridiaalisesti ja toinen ekvatoriaalisesti! Tällaista jakautumista kutsutaan ROTATIONAALISEKSI HOLOBLASTISEKSI HAJAUTUMISEKSI.
Nisäkkäiden jakautumisen toinen ainutlaatuinen piirre on, että blastomeerien jakautuminen on asynkronista. (verrattuna merisiilin ja sammakon synkronisuuteen aina keskiblastulan siirtymään asti). Nisäkkäiden alkion pilkkoutumista säätelee alusta alkaen zioottinen ydin.
Kolmannessa pilkkoutumisvaiheessa blastomeerit muodostavat löyhästi toisiinsa liittyneiden solujen pallon aivan kuten muutkin tutkimamme eläimet. Ennen neljättä pilkkoutumista blastulan solut muuttavat dramaattisesti käyttäytymistään toisiaan kohtaan. Ne pyrkivät nopeasti maksimoimaan kontaktinsa toisiin blastomeereihin ja saavat näin aikaan blastulan tiivistymisen.

Tämä KOKONAISUUS johtuu osittain uudenlaisen adheesiomolekyylin UVOMORULIININ (E-kadheriiniin) tuotannosta, ja sitä vakauttaa uloimpien solujen välisten tiukkojen yhteyksien muodostuminen, joka, kuten merisiilissä, eristää blastulan sisätilan ulkopuolelta. Solut muodostavat keskenään myös aukkoliitoksia, jotka mahdollistavat pienten molekyylien, kuten ionien ja joidenkin viestimolekyylien, kuten Ca++:n ja C-AMP:n, kulun. Tiivistetty 16-soluinen morula koostuu solujen ulkokuoresta ja muutamasta solusta (1-2), jotka ovat täysin sisäpuolella. Suurin osa ulkoisista soluista synnyttää TROBLASTISET TAI TROPHEKTODERMISET SOLUT. Nämä solut eivät osallistu varsinaiseen alkioon, vaan ne ovat sen sijaan välttämättömiä alkion istuttamiseksi kohdun seinämään, ja ne muodostavat KORIAN kudokset, joka on istukan olennainen osa, josta puhumme myöhemmin.
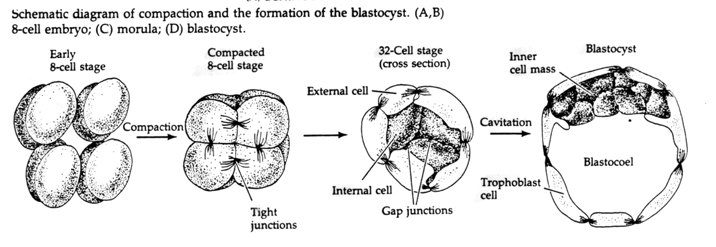
Alkiosolut ovat peräisin 16-soluisen blastulavaiheen sisemmistä muutamista soluista. Näistä soluista syntyy se sisempi solulimassa, josta koko alkio kehittyy. Kuudenteen halkioon eli 64 solun vaiheeseen mennessä sisäinen solumassa ja trofoblastikerros ovat täysin erillään. Trofoblastit erittävät nestettä sisäisesti luodakseen blastokotelon. Alkiota kutsutaan nyt blastokystaksi.
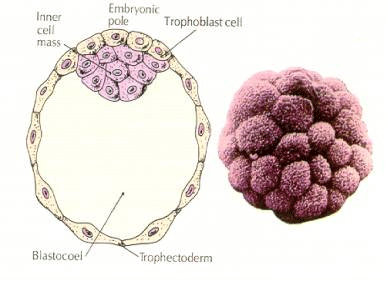 SISÄISEN SOLUMASSAN MUODOSTUMINEN
SISÄISEN SOLUMASSAN MUODOSTUMINEN
Miten nämä sisäisen solumassan solut syntyvät? Onko olemassa tiettyjä blastomeerejä, joiden kohtalona on sisäisten tekijöiden perusteella tulla sisäisen solumassan esiasteiksi? Vastaus näyttää olevan ei. Kaikki varhaiset blastomeerit näyttävät olevan totipotentteja, ja sen määrittäminen, mitkä solut muodostavat trofoblastikerroksen ja mitkä sisemmän solumassan, on vain sattumanvarainen kysymys. 4-soluisen alkion solut, jotka normaalisti tuottavat sekä sisäistä solumassaa että trofektodermisoluja, siirretään 32-soluisen alkion ulkopuolelle ja niistä syntyy vain trofektodermisoluja. Ne eivät vaikuta varsinaiseen alkioon. Muistetaan aiemmalta kloonausta käsittelevältä luennolta, että kahden 8 solun vaiheessa olevan hiiren alkion fuusio johtaa normaaliin alkioon, mikä viittaa siihen, että kaikki solut kyseisessä vaiheessa ovat totipotentteja.
MEROBLASTINEN HAJAUTUMINEN
Telo- ja sentroliitti- ja sentroliitti-alkion munasoluissa suuri tiheä keltuainen estää hajautumisen. Telolecithal-munat ovat tyypillisiä linnuille, kaloille ja matelijoille, kun taas centrolecithal-munat ovat tyypillisiä hyönteisille. Telolecithal-munat johtavat meroblastiseen diskoidiseen pilkkoutumiseen. Jakautuminen rajoittuu blastodiskiin munan eläinpuolella. Varhaisessa pilkkoutumisvaiheessa, koska pilkkoutuminen ei voi edetä keltarauhasen läpi, blastomeerit ovat kasvirajoiltaan yhtenäisiä.
Rolf Karlströmin elokuva seeprakalan kehityksestä on erinomainen. (Movie by Paul Myers)
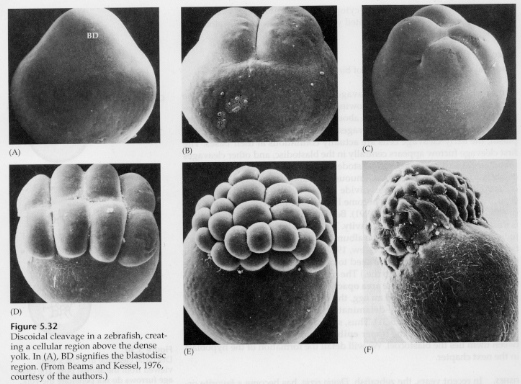 Vasta ekvatoriaalisissa halkeamisvaiheissa blastodermin solut irtoavat keltarauhasesta. Jatkuvat ekvaattorihalkeamat luovat monikerroksisen, kolme tai neljä solua paksun blastodermin.
Vasta ekvatoriaalisissa halkeamisvaiheissa blastodermin solut irtoavat keltarauhasesta. Jatkuvat ekvaattorihalkeamat luovat monikerroksisen, kolme tai neljä solua paksun blastodermin.
Linnuille muodostuu blastodermin ja keltarauhasen väliin tila, jota kutsutaan SUBGERMINAL-onteloksi. Blastodermin solut vaeltavat 16:nteen jakautumiseen mennessä (60 000 solua) subgerminaaliseen onteloon muodostaen toisen kerroksen. Näitä kahta kerrosta kutsutaan ulommaksi EPIBLASTIKSI ja sisemmäksi HYPOBLASTIKSI, joiden välissä on blastokotelo. Tutkimme tätä yksityiskohtaisemmin myöhemmin, kun käsittelemme lintujen ja nisäkkäiden gastrulaatiota
Niveljalkaisten niveljalkaisten munat käyvät läpi SUPERFIKTIIVISEN PUHDISTUKSEN. Suuri keltuaista sisältävä keskusmassa rajoittaa pilkkoutumisen munan sytoplasmiselle reunalle.
Hyönteisillä on mielenkiintoinen ja informatiivinen variaatio. Siellä zygoottiytimet jakautuvat ilman pilkkoutumista. Toisin sanoen ytimissä tapahtuu karyokinesis—-mitoottinen tuman jakautuminen— ilman sytokinesistä—solun jakautumista. Näitä alastomia tumia kutsutaan nimellä ENERGIDIT. Ytimet jakautuvat hämmästyttävällä nopeudella — joka 8. minuutti (koko alkionmuodostus kestää vain 22 tuntia).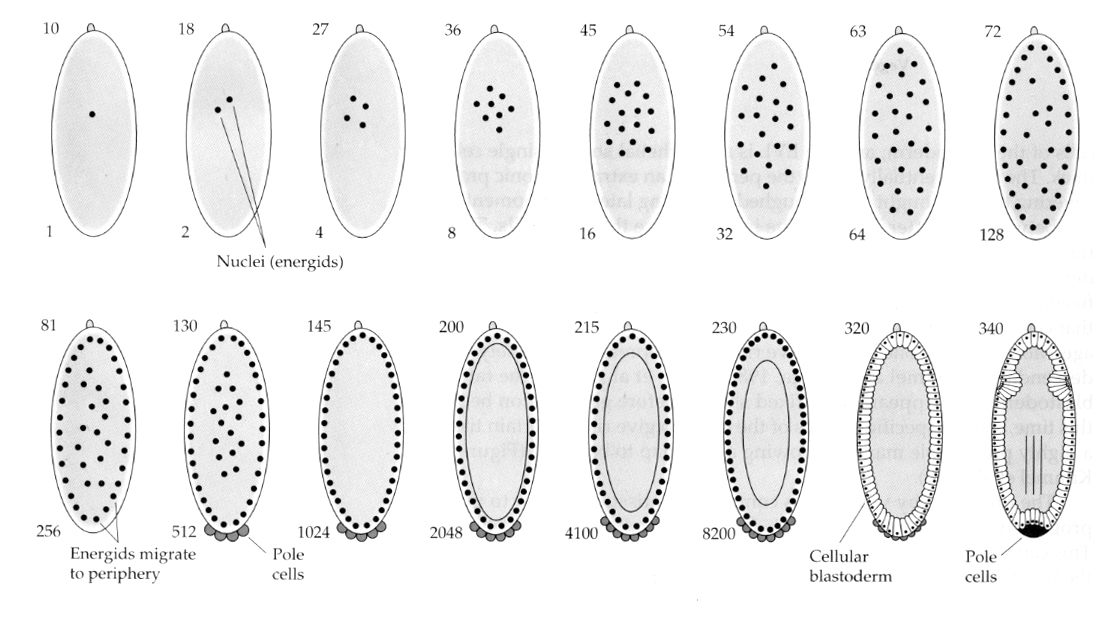
 Monien karyokinesiakierrosten jälkeen alastomat ytimet vaeltavat munasolun periferiaan. Tätä vaihetta kutsutaan SYNKYYTTISEKSI BLASTODERMIKSI, koska kaikki ytimet jakavat saman sytoplasman. Solunjakautuminen tapahtuu noin 14. ydinjakautumisen yhteydessä, jolloin syntyy SELLULAARINEN BLASTODERMI. Tämän jälkeen solut jakautuvat epäsynkronisesti. Tämä vastaa sammakoiden ja merisiilien midblastula-siirtymää. (siirtyminen maternaalisesta geeniekspressiosta pääasiassa zygoottiseen geeniekspressioon) Muistakaa, että midblastula-siirtymän ajateltiin käynnistyvän kromatiinin ja sytoplasman suhteen perusteella. Todisteet tästä mekanismista kärpäsillä nähdään tutkimalla mutantteja haploideja alkioita. Nämä alkiot käyvät läpi midblastula-siirtymän ja solunmuodostuksen yhtä jakautumista 15. myöhemmin. Lisäksi soluuntumista voidaan nopeuttaa sitomalla munasolu ja vähentämällä sytoplasman määrää. Vaikka synnynnäinen blastodermivaihe viittaa siihen, että kaikki ytimet ovat ekvipotentteja, koska sytoplasman määritteille ei näytä olevan diffuusioesteitä, sytoplasma on itse asiassa hyvin alueellistunutta, ja ytimillä on niiden ympärillä hyvin järjestäytyneitä sytoplasmallisia alueita.
Monien karyokinesiakierrosten jälkeen alastomat ytimet vaeltavat munasolun periferiaan. Tätä vaihetta kutsutaan SYNKYYTTISEKSI BLASTODERMIKSI, koska kaikki ytimet jakavat saman sytoplasman. Solunjakautuminen tapahtuu noin 14. ydinjakautumisen yhteydessä, jolloin syntyy SELLULAARINEN BLASTODERMI. Tämän jälkeen solut jakautuvat epäsynkronisesti. Tämä vastaa sammakoiden ja merisiilien midblastula-siirtymää. (siirtyminen maternaalisesta geeniekspressiosta pääasiassa zygoottiseen geeniekspressioon) Muistakaa, että midblastula-siirtymän ajateltiin käynnistyvän kromatiinin ja sytoplasman suhteen perusteella. Todisteet tästä mekanismista kärpäsillä nähdään tutkimalla mutantteja haploideja alkioita. Nämä alkiot käyvät läpi midblastula-siirtymän ja solunmuodostuksen yhtä jakautumista 15. myöhemmin. Lisäksi soluuntumista voidaan nopeuttaa sitomalla munasolu ja vähentämällä sytoplasman määrää. Vaikka synnynnäinen blastodermivaihe viittaa siihen, että kaikki ytimet ovat ekvipotentteja, koska sytoplasman määritteille ei näytä olevan diffuusioesteitä, sytoplasma on itse asiassa hyvin alueellistunutta, ja ytimillä on niiden ympärillä hyvin järjestäytyneitä sytoplasmallisia alueita.
KLEAVAATION MEKANISMIT
Solusykli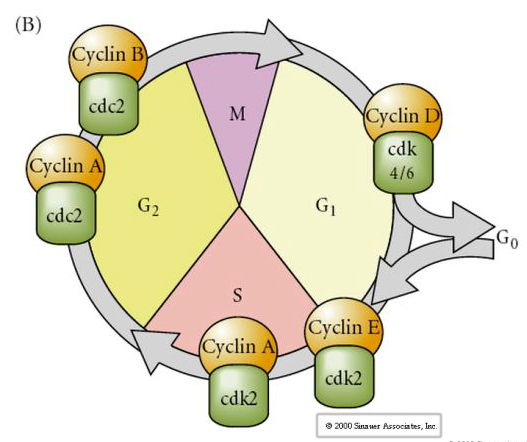
M-mitoosi
G1- pre-replikaatioväli
S- DNA-synteesi
G2-premitoottinen välikappale
Kleavage-vaiheen alkioissa, kuten sammakoissa ja kärpäsissä, blastomeerit kulkevat suoraan M:stä S:ään ilman G1- tai G2-vaiheita. Keskiblastomeren siirtymän jälkeen soluilla on molemmissa eläimissä G1- ja G2-vaihe. Elegantit siirtokokeet ovat osoittaneet, että sytoplasma säätelee sekä karyokinesistä että sytokinesistä. Jos jakautuvista soluista peräisin olevia tumia siirretään munasoluun, ne lopettavat välittömästi jakautumisen.
 Kääntäen, jos jakautumattomista soluista peräisin olevia tumia siirretään hedelmöittyneisiin enukleoituneisiin munasoluihin, ne alkavat jakautua. Keinotekoisesti aktivoiduissa enukleoituneissa munasoluissa, joissa ei ole sentrioleja, tapahtuu halkeilua muistuttavia kortikaalisia supistuksia. Joitakin solunjakautumista sääteleviä sytoplasmatekijöitä varhaisessa alkionvaiheessa on tunnistettu.
Kääntäen, jos jakautumattomista soluista peräisin olevia tumia siirretään hedelmöittyneisiin enukleoituneisiin munasoluihin, ne alkavat jakautua. Keinotekoisesti aktivoiduissa enukleoituneissa munasoluissa, joissa ei ole sentrioleja, tapahtuu halkeilua muistuttavia kortikaalisia supistuksia. Joitakin solunjakautumista sääteleviä sytoplasmatekijöitä varhaisessa alkionvaiheessa on tunnistettu.
SYTOSTAATTINEN FAKTORI (CSF) kohoaa ensimmäisen meioottisen jakautumisen jälkeen ja pysäyttää munasolun toisessa meioottisessa metafaasissa. Hedelmöitymisen yhteydessä Ca inaktivoi CSF:n, meioosi saadaan päätökseen ja pronominit fuusioituvat.
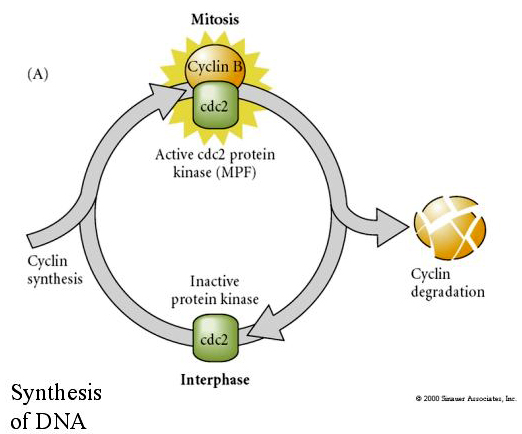
MITOSIS PROMOTING FACTOR (MPF) saa solut siirtymään M-vaiheeseen. MPF:n aktivoituminen aiheuttaa: 1. kromosomien tiivistymisen H1-histonin fosforylaatiolla, 2. ydinkuoren hajoamisen 3 ydinlamiinin hyperfosforylaatiolla, 3. RNA-polymeraasin eston transkription pysäyttämiseksi, 4. myosiinin säätelyalayksikön fosforylaation sytokinesiksen inhibtaatioksi.
Esitetty malli solusyklin sykliselle säätelylle embriksen pilkkoutumisvaiheiden aikana. MPF indusoi solun etenemään S:stä M:ään. CSF sitoutuu MPF:ään ja estää sen inaktivoitumisen. Solu pysyy M:ssä. Ca nousee ja aiheuttaa CSF:n inaktivoitumisen, mikä puolestaan johtaa MPF:n inaktivoitumiseen ja solu etenee M:n kautta S:ään ja sykli toistuu. MPF koostuu kahdesta alayksiköstä, sykliini B:stä ja cdc2:sta. Sykliini B syntetisoituu ja hajoaa solusyklispesifisesti solun ytimen säätelemänä, mikä ohjaa solusykliä normaaleissa somaattisissa soluissa. Oogeneesin aikana munasolu on kuitenkin täynnä sykliini B:n ”säätelijöitä” ja sykliini B:n mRNA:ta, joten sen synteesiä säätelevät zygootin ytimestä riippumattomat äidin tekijät. Näin ollen vasta kun äidin komponentit ”loppuvat”, zygoottinen ydin ottaa ohjat käsiinsä ja normaali solusykli (M, G1, S, G2) palaa takaisin.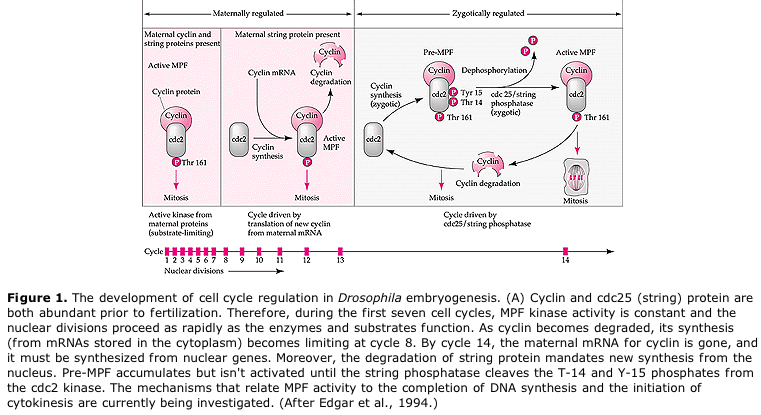
SELLUNKÄYTÖN MÄÄRITTÄMINEN
DETERMINANTTIEN sytoplasminen lokalisaatio yleisenä ja perustavanlaatuisena mekanismina varhaiseen kuviointikehitykseen (Esimerkit: Tunicate ja Sea Urchin). Kehitysbiologian keskeinen kysymys on, milloin ja miten solun kohtalot määräytyvät kehityksen aikana. Tämä liittyy läheisesti kysymykseen siitä, miten kuvionmuodostus tapahtuu kehityksen aikana. Alkion on paitsi tuotettava oikea määrä ja tyyppi erilaistuneita soluja myös järjestettävä ne oikealla tavalla suhteessa alkion kaikkiin muihin soluihin, jotta siitä muodostuisi toimiva eläin. Tarkastelemme kahta mahdollisuutta solujen kohtalon määräytymiseen ja kuvioiden muodostumiseen: 1. Solujen kohtalo voisi määräytyä sisäisten tekijöiden perusteella, jotka sijoitetaan munasoluun oogeneesin aikana ja jotka sitten jakautuvat tiettyihin blastomeereihin pilkkoutumisen aikana, 2. Alkion ympäristön antamat ulkoiset signaalit saattaisivat antaa solujen kohtaloa säätelevää kuviotietoa. Kuten tulemme näkemään, useimmat monimutkaiset organismit käyttävät sisäisten ja ulkoisten signaalien yhdistelmää säätelemään solujen kohtaloa ja alkion kuvionmuodostusta.
Autonominen solun kohtalon spesifikaatio sytoplasman determinanttien avulla viittaa siihen, että solun kohtalo on täysin riippuvainen sen sukupuolesta, kun taas ”säätelykehitys” viittaa siihen, että solun kohtalo määräytyy muilta soluilta tulevien ulkoisten signaalien mukaan. Nämä kaksi solujen spesifikaatiomekanismia voidaan erottaa toisistaan kokeellisesti eristämis-, ablaatio- ja siirtokokeilla. Jos alkiosta eristetty blastmere erilaistuu normaalisti (ikään kuin se olisi edelleen normaalissa paikassaan alkiossa), voimme sanoa, että sillä on oltava sisäisiä determinantteja, jotka määrittävät sen kohtalon. Jos se kuitenkin erilaistuu epänormaalisti, voimme sanoa, että sen solujen kohtalo on riippuvainen ulkoisista signaaleista. Jos poistamme alkion blastomeerin ja alkio kehittyy epänormaalisti, koska siitä puuttuvat kaikki solujen kohtalot, jotka normaalisti syntyvät poistetusta blastomeeristä, sanomme, että kehitys on solujen autonomista ja sisäisesti määriteltyä. Jos alkio kuitenkin kehittyy normaalisti, sanomme, että jäljellä olevat blastomeerit voivat säädellä solujensa kohtaloa kompensoidakseen puuttuvat solut. Jos siirretty solu säilyttää solun kohtalonsa alkuperäisen sijaintinsa perusteella, sanomme, että sen kohtalo on määräytynyt, jos taas se omaksuu uuden kohtalon juuri siirretyn sijaintinsa perusteella, sanomme, että sen kohtaloa säätelevät ulkoiset signaalit läheisiltä soluilta.
SYTOPLASMINEN SIJAINTI JA SÄÄTELY TUNIKAATTIMUNA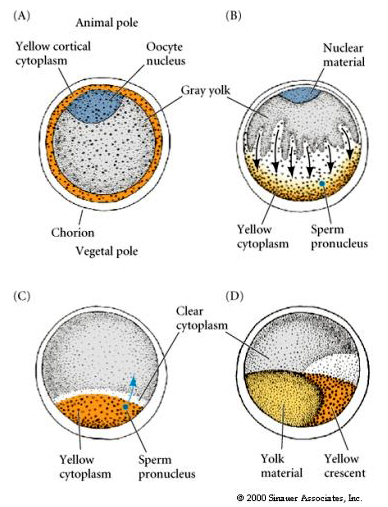
Oogeneesin lopussa tunikaattimunan eläin- ja kasvinapa erottuvat selvästi toisistaan. Munassa on keltainen kortikaalinen sytoplasma, joka ympäröi harmaata keltamaista sisempää sytoplasmaa. Munasolun tuma on siirtynyt kohti eläimellistä napaa. Siemennesteen tunkeutuminen kasvipuoliskoon hedelmöittää munasolun ja käynnistää kehityksen. Hedelmöityksen jälkeen munasolun sytoplasmassa tapahtuu dramaattinen uudelleenjärjestely, joka synnyttää alueellisesti värillisiä sytoplasmoja, jotka näyttävät korreloivan myöhempien blastomereiden kohtaloiden kanssa.
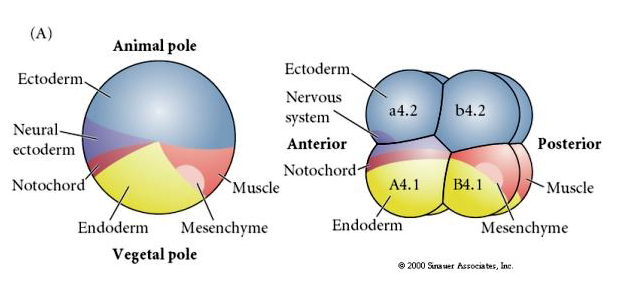 Huomaa, että kohtalokartta korreloi tunikaattialkion eriväristen sytoplasmien kanssa. Älä hämmenny kahdessa kuvassa olevista eri väreistä. ”Oranssi” keltainen sirppi-sytoplasma korreloi lihasten kohtaloiden kanssa ja yolky (keltainen) sytoplasma korreloi endodermien kohtaloiden kanssa. Harmaa (valkoinen tai sinertävän violetti) sytoplasma keltaisen puolikuun yläpuolella korreloi neuraalisen ektodermin kanssa.
Huomaa, että kohtalokartta korreloi tunikaattialkion eriväristen sytoplasmien kanssa. Älä hämmenny kahdessa kuvassa olevista eri väreistä. ”Oranssi” keltainen sirppi-sytoplasma korreloi lihasten kohtaloiden kanssa ja yolky (keltainen) sytoplasma korreloi endodermien kohtaloiden kanssa. Harmaa (valkoinen tai sinertävän violetti) sytoplasma keltaisen puolikuun yläpuolella korreloi neuraalisen ektodermin kanssa.
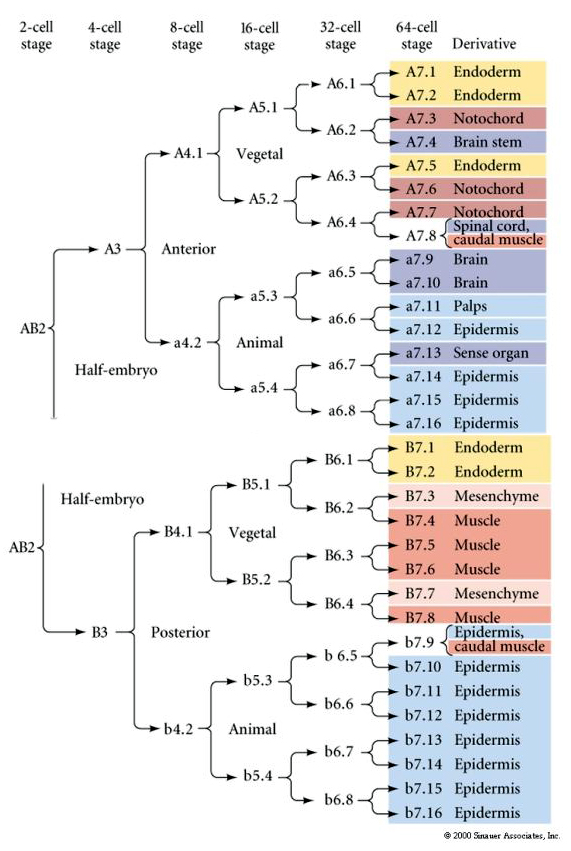 Tässä linjakartassa näkyy invarianttien linjojen korrelaatio, jossa blastomeerit on paketoitu tiettyihin värillisiin sytoplasmiin invarianttien soluhalkeamien mukaan. Invariantit pilkkoutumat ja linjalinjat eivät kuitenkaan välttämättä todista solujen autonomista spesifikaatiota sytoplasman determinanttien avulla.
Tässä linjakartassa näkyy invarianttien linjojen korrelaatio, jossa blastomeerit on paketoitu tiettyihin värillisiin sytoplasmiin invarianttien soluhalkeamien mukaan. Invariantit pilkkoutumat ja linjalinjat eivät kuitenkaan välttämättä todista solujen autonomista spesifikaatiota sytoplasman determinanttien avulla.
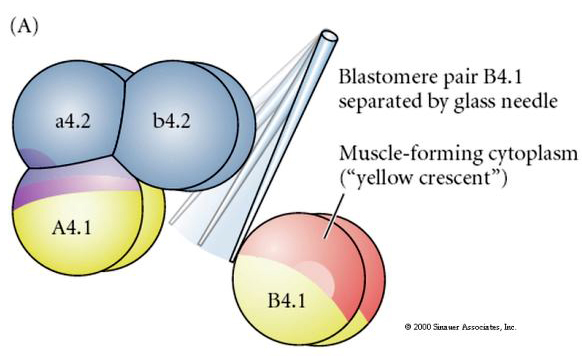 Tarvitaan kokeellisia manipulaatioita, jotta voidaan testata regulatiivista vs. solujen autonomista solun kohtalon määrittelyä. Seuraavissa kolmessa kuvassa esitetyillä klassisilla eristyskokeilla pyritään osoittamaan, että solujen kohtalo määräytyy sytoplasman determinanttien perusteella, jotka ne saavat stereotyyppisten pilkkomisten kautta. Lasineulaa käytetään B4.1-blastomeeriparin erottamiseen muusta alkion osasta. B4.1-blastomeerit saavat normaalisti lihassolujen kohtaloon korreloituvan keltaisen crecent-sytoplasman.
Tarvitaan kokeellisia manipulaatioita, jotta voidaan testata regulatiivista vs. solujen autonomista solun kohtalon määrittelyä. Seuraavissa kolmessa kuvassa esitetyillä klassisilla eristyskokeilla pyritään osoittamaan, että solujen kohtalo määräytyy sytoplasman determinanttien perusteella, jotka ne saavat stereotyyppisten pilkkomisten kautta. Lasineulaa käytetään B4.1-blastomeeriparin erottamiseen muusta alkion osasta. B4.1-blastomeerit saavat normaalisti lihassolujen kohtaloon korreloituvan keltaisen crecent-sytoplasman.
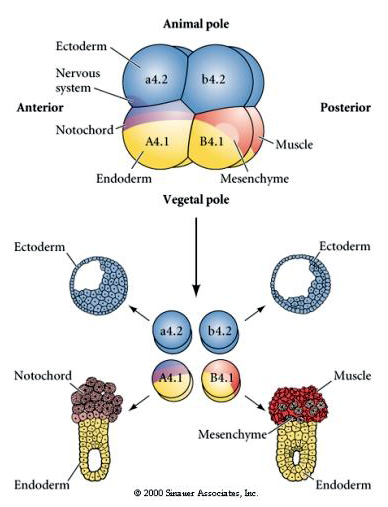
Tässä nähdään eristyskokeiden tulokset. Kussakin tapauksessa eristetyistä blastomeereistä syntyy vain se osajoukko solupatoja, joita ne normaalisti tuottaisivat ehjässä alkiossa. Eristetyt blastomeerit eivät säädä kohtaloaan kompensoidakseen puuttuvia naapureitaan. Eläinten napojen blastomeerit, a4.2 ja b4.2, synnyttävät vain ektodermisoluja. A4.1:stä syntyy notokordia ja endodermisoluja, kun taas B4.1:stä syntyy lihas- ja endodermisoluja. Yksikään eristetyistä blastomeereistä ei voi synnyttää kaikkia normaalin alkion solukomponentteja.
Seuraavassa kokeessa manipuloidaan neulalla ekvatoriaalista pilkkoutumistasoa niin, että se on normaalia kasvullisempi, ja nyt eläimen napaan kuuluvat blastomeerit, b4.2, saavat osan ”keltaisen puolikuun” sytoplasmasta. Kun nämä blastomeerit eristetään, niistä syntyy nyt joitakin lihassoluja. Tämä osoittaa hienosti, että ”keltainen puolikuu” -sytoplasma voi määrittää lihassolujen kohtalon ja voi tehdä sen soluautonomisesti.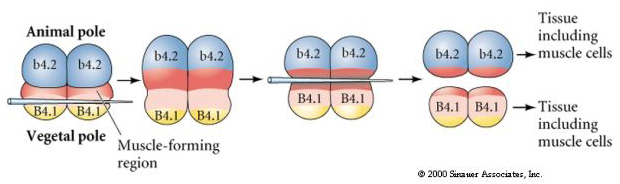
LOKALISAATIO JA SÄÄTELY MERIUURINMUNASSA
Hyytelökanava määrittelee eläinnavan sijainnin ja kuvastaa munan varhaista polariteettia. Halkioiden varhainen malli ei riipu siittiöiden sisäänmenopaikasta, vaan ne määräytyvät munasolun luontaisen polariteetin/asymmetrian mukaan. Boveri (1901) kuvasi subequatoriaalisen pigmenttikaistaleen, joka on järjestetty ortongonaalisesti eläin-vegetaaliakseliin nähden. Nämä rakeet osoittivat myös sytoplasman sijainnin, joka myöhemmin sisältyy arkkenteronin soluihin. Horstadius (1928) erotti eläin- ja kasviperäiset blastomeerit toisistaan ja osoitti, että vain kasviperäinen blastomeeri synnyttää mikromerejä, gastruloi ja muodostaa luuston. Hänen johtopäätöksensä oli, että kasvipuolikkaassa sijaitsevat sytoplasmatekijät ovat välttämättömiä mikromereille, gastrulaatiolle ja arkkenteronin muodostumiselle sekä luurangon muodostumiselle.Muistakaa varhaisten halkioiden malli. Mikromeerit syntyvät neljännen pilkkoutumisen aikana (16 solun vaiheessa) kasvullisen puolen blastomeerien epätasaisesta ekvatoriaalisesta jakautumisesta.
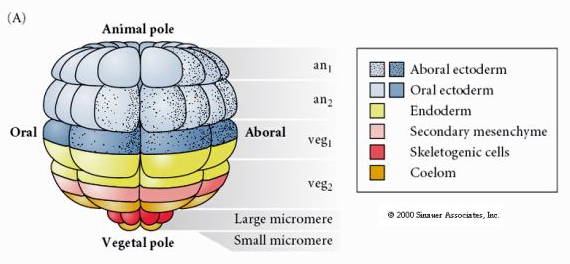 Tässä näkyy 64 solun vaiheessa olevan merisiilin blastulan kohtalokartta. Huomaa, että mikromerit ovat primaarisia mesenkyymisoluja, ja niistä syntyy toukkien luuranko (pluteusvaiheen piikit).
Tässä näkyy 64 solun vaiheessa olevan merisiilin blastulan kohtalokartta. Huomaa, että mikromerit ovat primaarisia mesenkyymisoluja, ja niistä syntyy toukkien luuranko (pluteusvaiheen piikit).
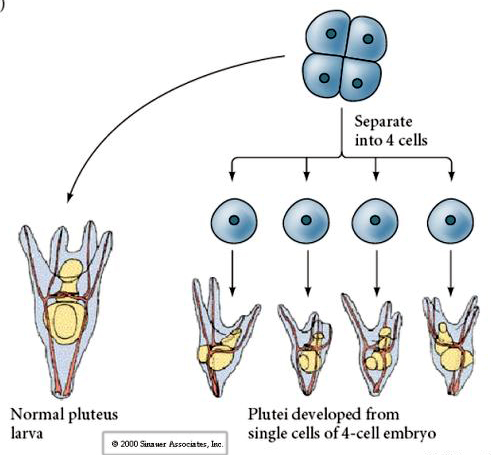 Neljäsoluvaiheessa, jos blastomeerit eristetään toisistaan, ne pystyvät ”säätelemään” kohtaloaan ja synnyttämään 4 pientä pluteusvaiheen toukkaa.
Neljäsoluvaiheessa, jos blastomeerit eristetään toisistaan, ne pystyvät ”säätelemään” kohtaloaan ja synnyttämään 4 pientä pluteusvaiheen toukkaa.
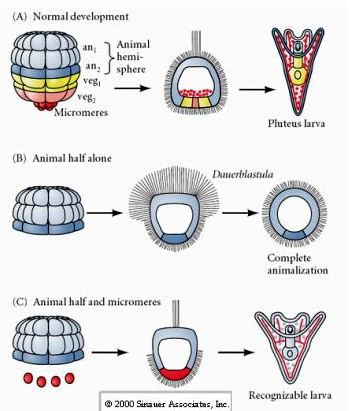 Sitä vastoin myöhemmissä vaiheissa, jos eläimelliset puolikkaat blastomeerit eristetään, havaitaan, että ne tuottavat vain ”eläimellistyneen” dauerblastulan, joka ei ilmentä mitään mesodermi- tai endodermisolujen kohtaloita. Eristetyt kasviperäiset puoliblastomeerit synnyttävät toukkia, jotka ilmentävät ektodermi-, mesodermi- ja endodermisolujen kohtaloita, mikä osoittaa, että näiden solujen kohtaloa voidaan säädellä. Eristetyt mikromereemit (primaarinen mesenkyymi) käyvät läpi oikean määrän solunjakautumia ja synnyttävät AINA aikataulun mukaisesti piikkejä. Näin ollen mikromeerit ovat lopullisesti määriteltyjä luuston muodostavien mesenkiimisolujen esiasteiksi, kun ne ilmestyvät ensimmäisen kerran 16 solun vaiheessa. Keskeisiä kokeita olivat mikromereiden yhdistäminen eläinten napojen blastomeereihin ja sen osoittaminen, että vaikka mikromereiden kohtalo oli ”kiinteä tai määräytynyt” niiden syntyhetkellä, mikromereillä oli kyky ”indusoida” uusia solujen kohtaloita eläinten napojen blastomeereihin. Mikromereerit pystyivät indusoimaan endodermisen ja mesodermisen kohtalon eläinnavan blastomeereihin! Näin ollen C-kohdan myöhäinen koe osoittaa, että kun mikromeerejä lisätään eläinpuolen blastomereihin, voidaan nyt indusoida tunnistettavan toukan muodostuminen, joka ilmentää endodermi-, mesodermi- ja ektodermi-patteja.
Sitä vastoin myöhemmissä vaiheissa, jos eläimelliset puolikkaat blastomeerit eristetään, havaitaan, että ne tuottavat vain ”eläimellistyneen” dauerblastulan, joka ei ilmentä mitään mesodermi- tai endodermisolujen kohtaloita. Eristetyt kasviperäiset puoliblastomeerit synnyttävät toukkia, jotka ilmentävät ektodermi-, mesodermi- ja endodermisolujen kohtaloita, mikä osoittaa, että näiden solujen kohtaloa voidaan säädellä. Eristetyt mikromereemit (primaarinen mesenkyymi) käyvät läpi oikean määrän solunjakautumia ja synnyttävät AINA aikataulun mukaisesti piikkejä. Näin ollen mikromeerit ovat lopullisesti määriteltyjä luuston muodostavien mesenkiimisolujen esiasteiksi, kun ne ilmestyvät ensimmäisen kerran 16 solun vaiheessa. Keskeisiä kokeita olivat mikromereiden yhdistäminen eläinten napojen blastomeereihin ja sen osoittaminen, että vaikka mikromereiden kohtalo oli ”kiinteä tai määräytynyt” niiden syntyhetkellä, mikromereillä oli kyky ”indusoida” uusia solujen kohtaloita eläinten napojen blastomeereihin. Mikromereerit pystyivät indusoimaan endodermisen ja mesodermisen kohtalon eläinnavan blastomeereihin! Näin ollen C-kohdan myöhäinen koe osoittaa, että kun mikromeerejä lisätään eläinpuolen blastomereihin, voidaan nyt indusoida tunnistettavan toukan muodostuminen, joka ilmentää endodermi-, mesodermi- ja ektodermi-patteja.
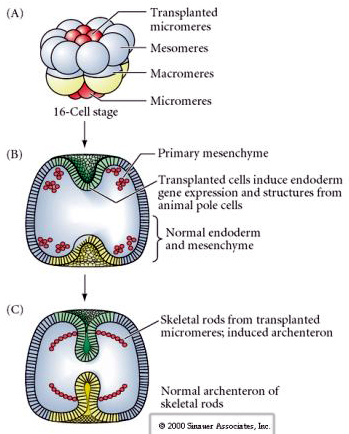 Viimeinen koesarja osoittaa, että jopa normaalissa alkiossa, jos mikromeerejä siirretään eläinpuolen korkkiin, voidaan indusoida sekundaarinen arkkenteroni ja muuttaa normaalia aksiaalista kuviointia. Tämä taas puoltaa sitä, että mikromereenit hankkivat sytoplasman derminantin, joka määrittää niiden solun kohtalon, ja että ne antavat induktiivisen signaalin, joka kuvioi merikihokki-alkion aksiaaliset rakenteet. Mikromereiden kohtaloa ei voida muuttaa, mutta mikromereistä tulevat signaalit voivat muuttaa kaikkien muiden blastomeerien kohtaloa.
Viimeinen koesarja osoittaa, että jopa normaalissa alkiossa, jos mikromeerejä siirretään eläinpuolen korkkiin, voidaan indusoida sekundaarinen arkkenteroni ja muuttaa normaalia aksiaalista kuviointia. Tämä taas puoltaa sitä, että mikromereenit hankkivat sytoplasman derminantin, joka määrittää niiden solun kohtalon, ja että ne antavat induktiivisen signaalin, joka kuvioi merikihokki-alkion aksiaaliset rakenteet. Mikromereiden kohtaloa ei voida muuttaa, mutta mikromereistä tulevat signaalit voivat muuttaa kaikkien muiden blastomeerien kohtaloa.
Horstadius: (1928, 1935) osoitti kokeellisesti, että 16 solun vaiheessa olevassa alkiossa kaikki blastomeerien kerrokset mikromerejä lukuun ottamatta saavat erilaisen kohtalon, kun ne siirretään eri paikkoihin kimeerisissä alkioissa. Archenteron kehittyy veg 1:n blastomeereistä, jos veg 2:n solut poistetaan ja mikromeerit asetetaan kosketuksiin veg 1:n kerroksen kanssa. Jos mikromerejä ei ole, veg 2 -blastomeereistä syntyy archenteronia ja luurankorakenteita. Klassisesti näiden tulosten selittämiseksi on vedottu kaksoiseläin-kasviperäiseen gradienttiin. Nämä tulokset viittaavat kuitenkin vain siihen, että vierekkäisten blastomeerikerrosten välillä esiintyy ratkaisevia induktiivisia vuorovaikutuksia.
Implantoiduilla yksittäisillä mikromeereillä lähellä eläimen napaa estetään apikaalisten tuppien muodostumista ja joissakin tapauksissa indusoidaan uusia alkion akseleita. Veg 2 -blastomeerit aiheuttavat myös samanlaisia muutoksia kuin mikromereerit, kun ne siirretään eläinnavan blastomeerien viereen.
SIIRTOJEN YLEISET TULOKSET: Tietyn blastomeerin kohtaloon havaitaan aina vaikuttavan normaalissa alkiossa niiden vieressä olevien erilaisten naapurisolujen kiinnittyminen.
HYPOTEESI: Paikalliset äidin sytoplasman determinantit määrittelevät tietyt solut normaalissa alkiossa, erityisesti mikromereenit ja arhenteronien esiasteet lähellä vegetatiivista napaa. Nämä solut määräävät sitten induktiivisesti viereisten blastomeerien kohtalot, jotka puolestaan ovat vuorovaikutuksessa naapureidensa kanssa. Monet blastomeereistä säilyttävät muita kuin normaalisti ilmentämiään potentiaaleja, ja jonkin aikaa nämä blastomeerit ovat vain palautuvasti spesifioituneita, kuten vaaditaan kehitysjärjestelmässä, joka riippuu suurelta osin induktiosta.